Анатомия женских половых органов. Репродуктивная женская система: эмбриология, анатомия, органы
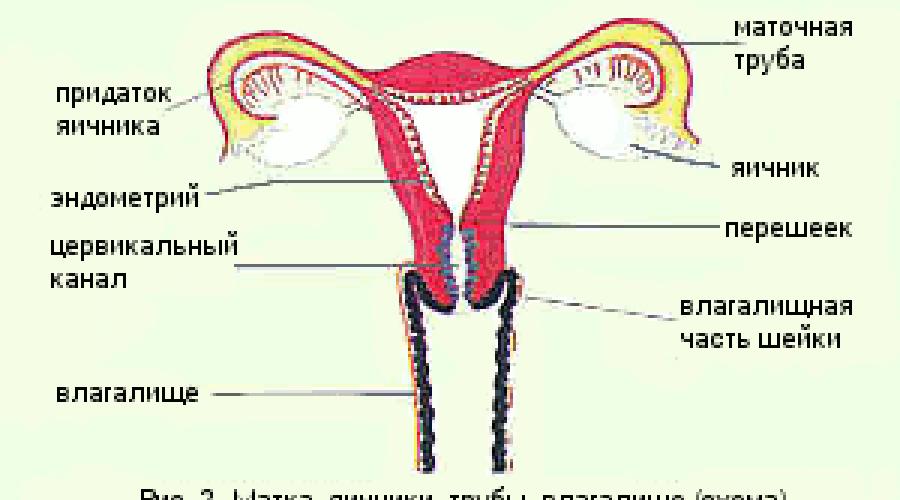
Форма матки грушевидная, уплощенная в переднезаднем направлении. От верхнебоковых краев матки отходят широкие маточные связки, в которых располагаются маточные (фаллопиевы) трубы и яичники (рис.1). Анатомически в матке различают дно, тело и шейку.
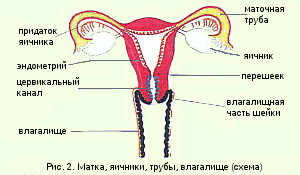
Дно – часть матки, расположенная выше отхождения маточных труб. Тело треугольной формы, суживается к перешейку. Полость матки также треугольной формы, в верхних углах расположены два отверстия, открывающиеся в маточные трубы, в нижнем углу расположен перешеек – сужение, ведущее в полость канала шейки матки (рис.2).
Шейка матки является относительно узким нижним сегментом матки. У девочек и девушек имеет коническую форму, у взрослой женщины цилиндрическую. Выделяют влагалищную часть (portio vaginalis cervicis - эктоцервикс), цервикальный канал (canalis cervicalis uteri – эндоцервикс) и перешеек. В шейке матки имеются два отверстия: внутренний зев – отверстие в верхнем отделе, расположенное на границе тела и шейки матки, и наружный зев – отверстие в нижнем отделе, открывающееся во влагалище.
Влагалищная часть шейки матки округлой формы, поверхность ее гладкая, в центре расположен наружный зев. У нерожавших женщин он маленький, округлой или поперечно-овальной формы (рот маленькой рыбки). После родов наружный зев приобретает форму поперечной щели. Канал шейки матки узкий, расширен в средней части. На передней и задней поверхности имеются два продольных гребня, от которых отходят под углом складки слизистой оболочки, пальмовидные складки. Эти образования придают каналу причудливый вид и названы деревом жизни.
Влагалище – мышечно-эластичная трубка, расположенная в малом тазу, верхней частью охватывает шейку матки, а нижним открывается в половую щель.
Слизистая оболочка тела матки состоит из стромы и однослойного цилиндрического эпителия, врастающего в строму с образованием простых трубчатых желез. Слизистая оболочка перешейка сходна со слизистой оболочкой тела матки и представлена большим числом клеток соединительной ткани и единичными простыми неветвящимися железами. Слизистая оболочка тела и перешейка претерпевает циклические изменения в течение менструального цикла.
Стенка шейки матки состоит преимущественно из коллагеновой ткани, строма слизистой оболочки содержит много эластических волокон. Железы эндоцервикса трубчатые, ветвящиеся, их не считают истинными железами, так как их строение на всем протяжении одинаково. Железы содержат секрет в виде густой стекловидной слизи, имеющей щелочную реакцию. Щелочная реакция способствует сохранению жизнеспособности сперматозоидов, их продвижению в полость матки. Секреция слизи в овуляцию увеличивается, секрет заполняет цервикальный канал и образует так называемую пробку Кристеллера, которая благодаря бактерицидным свойствам и механически препятствует проникновению в канал и полость матки микробов. Если железы закупориваются, а слизь продолжает накапливаться, образуются Наботовы кисты, которые могут выпячиваться на поверхность шейки.
Слизистая оболочка канала предствалена цилинрическим слизепродуцирующим эпителием, присутствуют единичные реснитчатые клетки, их число с возрастом значительно уменьшается.
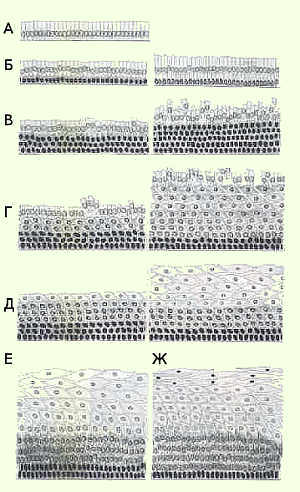
Слизистая оболочка эктоцервикса и влагалища выстланы многослойным плоским неороговевающим эпителием. В норме в репродуктивном возрасте эпителий состоит из множества рядов, условно разделенных на три слоя: базальный, промежуточный, поверхностный. Только нижний (базальный) слой клеток связан с базальной мембраной, клетки в нем располагаются в один ряд. Слой молодых клеток, расположенный над базальным, состоящий из нескольких рядов (нижняя часть промежуточного слоя). Называют парабазальным. Размер клеток по мере созревания увеличивается. Размер ядер уменьшается (рис.3)
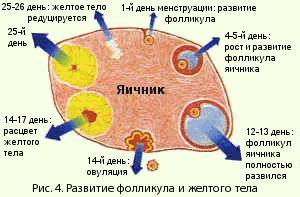
Строение многослойного плоского эпителия влагалища и влагалищной части шейки матки зависит от гормонального состояния женщины, фазы менструального цикла. В менструальном цикле различают 4 фазы: менструальную, фолликулиновую (эстрогенную, пролиферативную), овуляторную и лютеиновую (прогестиновую, секреторную). Эти фазы связаны с созреванием яйцеклетки, которое регулируется гонадотропными гормонами гипоталамо-гипофизарной системы (передней доли гипофиза). Под действием ФСГ в яичнике растет и созревает фолликул (рис.4).
Растущий фолликул выделяет эстрогеновые гормоны, определенное количество которых тормозит продукцию ФСГ и стимулирует выделение ЛГ. ЛГ совместно с ФСГ подготавливает фолликул к овуляции, а после разрыва фолликула (овуляции), если беременность не наступила, способствует превращению его в желтое тело.
Наиболее подвержен гормональным воздействиям эпителий влагалища, поэтому на изучение состава этого эпителия основаны гормональная цитологическая диагностика (см. Гормональная цитологическая диагностика).
Цилиндрический эпителий канала переходит в плоский в так называемой зоне стыка, расположенной у девочки на влагалищной порции шейки матки, у женщины репродуктивного возраста – на уровне наружного зева. Зона стыка под действием гормональных и других воздействий может перемещаться на влагалищную порцию шейки матки. Появление цилиндрического эпителия на влагалищной порции шейки называется эктопией. Под действием содержимого влагалища участок эктопии подвергается физиологическим изменениям, метаплазии в плоский эпителий (рис.6).
 |
1 – зрелый эпителий. На поверхности зрелые клетки поверхностного слоя с пикнотичными ядрами 2 – эпителий созревает до промежуточного слоя, на поверхности зрелые промежуточные клетки 3 – эпителий созревает до промежуточного слоя, на поверхности незрелые промежуточные клетки 4 - 5 - эпителий созревает только до парабазального слоя, на поверхности парабазальные клетки |
Рис.5. Разные стадии созревания многослойного плоского эпителия |
|
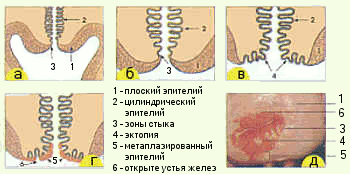 |
А- до полового созревания (зона стыка расположена в цервикальном канале) Б – В – во время полового созревания (зона стыка смещается на влагалищную часть шейки матки) Г – цилиндрический эпителий на влагалищной части замещается метаплазированным Д – кольпоскопическая картина шейки матки у многорожавшей женщины |
Рис.6. Расположение цилиндрического эпителия и зоны стыка в шейке матки |
|
Сформировавшееся желтое тело под действием ЛГ гипофиза выделяет прогестерон. Существует тесная взаимосвязь между гормонами яичника и надпочечника, яичника и щитовидной железы.
Эстрогены стимулируют полное созревание многослойного плоского неороговевающего эпителия до поверхностных клеток. Прогестерон тормозит созревание, и если его вырабатывается много, клетки созревают только до промежуточного слоя. В период постменопаузы, в связи со снижением выработки половых гормонов, эпителий подвергается атрофии (рис.5).
Цилиндрический эпителий на влагалищной части замещается метаплазированным. Участок метаплазированного эпителия носит название зоны трансформации, или зоны превращения. У женщин репродуктивного возраста зона стыка, как правило представлена участками естественной зоны стыка, участками зоны трансформации, метаплазированным эпителием. Зона трансформации может располагаться на влагалищной части шейки матки, а может (полностью или частично) уходить в цервикальный канал. У женщин в постменопаузе зона стыка и зона трансформации чаще всего располагается в цервикальном канале. Зона трансформации является наиболее опасной с точки зрения возможности патологических, в том числе неопластических изменений.
Метаплазированный плоский эпителий (рис.7) развивается не из зрелых цилиндрических, а из подцилиндрических, так называемых резервных клеток. В норме обычно резервные клетки не обнаруживаются в гистологических и цитологических препаратах. Резервноклеточная гиперплазия является первым этапом плоскоклеточной метаплазии. Под слоем цилиндрических клеток появляются один, два и более слоев клеток герминативного типа, которые напоминают клетки базального слоя плоского эпителия без четких клеточных границ.
Зрелый метаплазированный эпителий морфологически практически неотличим от "естественного" плоского, представлен всеми слоями, типичными для многослойного плоского неороговевающего эпителия.
 |
А - цилиндрический эпителий Б – под слоем цилиндрического эпителия появляется слой подцилиндрических (резервных) клеток В – резервные клетки размножаются, цилиндрические слущиваются с поверхности Г – этап незрелой плоскоклеточной метаплазии: определяются четкие границы резервных клеток и постепенно формируется 3-4 слоя клеток, сходных с многослойным плоским неороговевающим эпителием Д – этап созревающей плоскоклеточной метаплазии. На поверхности эпителиального пласта клетки средних размеров с небольшими ядрами Е - этап зрелой плоскоклеточной метаплазии. Клетки на поверхности эпителиального пласта сходны с промежуточными клетками плоского эпителия Ж - этап зрелой плоскоклеточной метаплазии. Клетки на поверхности эпителиального пласта практически неотличимы от "естественных" клеток поверхностного слоя плоского эпителия |
Рис.7. Этапы плоскоклеточной метаплазии |
|
(Документ)
n1.doc
ГЛАВА 2АНАТОМИЯ И ФИЗИОЛОГИЯ ЖЕНСКОЙ РЕПРОДУКТИВНОЙ СИСТЕМЫ
2.1. АНАТОМИЯ ЖЕНСКИХ ПОЛОВЫХ ОРГАНОВ
Женская половая система состоит из внутренних половых органов (внутренние гениталии), расположенных в полости малого таза, и наружных половых органов (наружные гениталии), находящихся вне костного таза. К внутренним половым органам относятся матка, яичники, маточные трубы и влагалище. Наружные половые органы включают лобок, большие и малые половые губы, преддверие влагалища и клитор. Полость таза ограничивается костными структурами. Дном для внутренних половых и смежных с ними органов служит диафрагма таза, состоящая из мышц и фасций.
2.1.1. Женский таз
Таз (см. раздел 2.1.3) состоит из четырех костей: двух тазовых (или безымянных), крестца и копчика.
Тазовая кость (os coxae) до 16-18 лет состоит из трех костей, соединенных хрящами: подвздошной, лобковой и седалищной. После окостенения хрящей указанные кости срастаются между собой, образуя тазовую (безымянную) кость.
Подвздошная кость (os ilium) состоит из двух частей: тела и крыла. Тело - это короткая утолщенная часть кости, участвующая в образовании вертлужной впадины. Крыло представляет собой широкую пластинку с вогнутой внутренней и выпуклой наружной поверхностью. Наиболее утолщенный свободный край крыла образует гребень подвздошной кости (crista iliaca). Спереди гребень начинается верхней передней подвздошной остью, или выступом (spina iliaca anterior superior), ниже располагается нижняя передняя ость (spina iliaca anterior inferior).
Гребень подвздошной кости сзади заканчивается верхней задней подвздошной остью (spina iliaca posterior superior), ниже которой располагается второй выступ - нижняя задняя подвздошная ость (spina iliaca posterior inferior). На внутренней поверхности подвздошной кости в области перехода крыла в тело располагается гребневый выступ, который образует дугообразную, или пограничную, или безымянную, линию (linea arcuata, s.terminalis, s.innominata). Эта линия идет от крестца поперек всей подвздошной кости, спереди переходят на верхний край лонной кости.
Седалищная кость (os ischii) состоит из тела, участвующего в образовании вертлужной впадины, и двух ветвей: верхней и нижней. Верхняя ветвь идет от тела книзу и заканчивается седалищным буфом (tuber ischiadicum). На задней поверхности нижней ветви имеется выступ - седалищная ость (spina ischiadica). Нижняя ветвь направляется кпереди и кверху и соединяется с нижней ветвью лонной кости.
Лобковая кость (os pubis) образует переднюю стенку таза. Она состоит из тела и двух ветвей: верхней (горизонтальной) и нижней (нисходящей). Короткое тело лобковой кости составляет часть вертлужной впадины, нижняя ветвь соединяется с соответствующей ветвью седалищной кости.
Верхние и нижние ветви обеих лобковых костей спереди соединяются друг с другом посредством малоподвижного лобкового сочленения - симфиза (symphisis). Обе лобковые кости соединяются в симфизе промежуточным хрящом, в котором нередко бывает маленькая щелевидная полость, заполненная жидкостью. Нижние ветви лобковых костей образуют под симфизом угол, который называется лобковой дугой. Соединяющие ветви лобковой и седалищной костей ограничивают довольно обширное запирательное отверстие (foramen obturatorium).
Крестец (os sacrum) состоит из пяти сросшихся позвонков. Величина крестцовых позвонков уменьшается по направлению книзу, поэтому крестец имеет форму усеченного конуса. Широкая его часть - основание крестца - обращена вверх, узкая часть - верхушка крестца - вниз. Задняя поверхность крестца выпуклая, передняя - вогнутая, она образует крестцовую впадину. Основание крестца (поверхность I крестцового позвонка) сочленяется с V поясничным позвонком; на середине передней поверхности основания крестца образуется выступ - крестцовый мыс (promontorium).
Копчик (os coccygis) состоит из 4-5 рудиментарных сросшихся копчиковых позвонков, представляет собой небольшую кость, суживающуюся книзу.
Кости таза соединяются посредством симфиза, крестцово-подвздошных и крестцово-копчикового сочленений. В сочленениях таза располагаются хрящевые прослойки. Сочленения таза укреплены прочными связками.
2.1.2. Наружные и внутренние половые органы
Наружные половые органы (genitalia externa, s. vulva), имеющие собирательное название "вульва", или "пудендум", находятся ниже лобкового симфиза (рис. 2.1). К ним относятся лобок, большие и малые половые губы, клитор и преддверие влагалища. В преддверие влагалища открываются наружное отверстие мочеиспускательного канала (уретра) и протоки больших желез преддверия (бартолиновы железы).
Лобок (mons pubis), пограничный участок брюшной стенки, представляет собой округлое срединное возвышение, лежащее спереди от лобкового симфиза и лобковых костей. После полового созревания он покрывается волосами, а его подкожная основа в результате интенсивного развития приобретает вид жировой подушечки.
Большие половые губы (labia pudendi majora) - широкие продольные складки кожи, содержащие большое количество жировой клетчатки и фиброзные окончания круглых маточных связок. Спереди подкожная жировая клетчатка большой половой губы переходит в жировую подушечку на лобке, а сзади соединена с седалищно-прямокишечной жировой клетчаткой. После достижения половой зрелости кожа наружной поверхности больших половых губ пигментируется и покрывается волосами. В коже больших половых губ находятся потовые и сальные железы. Внутренняя поверхность их гладкая, не покрыта волосами и насыщена сальными железами. Соединение больших половых губ спереди называют передней спайкой, сзади - спайкой половых губ, или задней спайкой. Узкое пространство перед задней спайкой половых губ называют ладьевидной ямкой.
Рис. 2.1. Наружные половые органы.
1 - лобок; 2 - передняя спайка; 3 - большая половая губа; 4 - малая половая губа; 5 - задняя стенка влагалища; 6 - ямка преддверия влагалища; 7 - задняя спайка (спайка половых губ); 8 - задний проход; 9 - промежность; 10 - вход во влагалище; 11-свободный край девственной плевы; 12 -наружное отверстие мочеиспускательного канала; 13 - уздечка клитора; 14 - клитор.
Малые половые губы (labia pudendi minora). Толстые складки кожи меньших размеров, называемые малыми половыми губами, находятся медиально от больших половых губ. В отличие от больших половых губ они не покрыты волосами и не содержат подкожной жировой клетчатки. Между ними находится преддверие влагалища, которое становится видимым только при разведении малых половых губ. Спереди, где малые половые губы подходят к клитору, они делятся на две небольшие складки, которые сливаются вокруг клитора. Верхние складки соединяются над клитором и образуют крайнюю плоть клитора; нижние складки соединяются на нижней стороне клитора и образуют уздечку клитора.
Клитор (clitoris) находится между передними концами малых половых губ под крайней плотью. Он является гомологом пещеристых тел мужского полового члена и способен к эрекции. Тело клитора состоит из двух пещеристых тел, заключенных в фиброзную оболочку. Каждое пещеристое тело начинается ножкой, прикрепленной к медиальному краю соответствующей седалищно-лобковой ветви. Клитор прикрепляется к лобковому симфизу с помощью поддерживающей связки. На свободном конце тела клитора находится небольшое возвышение эректильной ткани, которое называется головкой.
Луковицы преддверия (bulbi vestibuli) - венозные сплетения, расположенные в глубине малых половых губ и подковообразно охватывающие преддверие влагалища. Рядом с преддверием влагалища вдоль глубокой стороны каждой малой половой губы расположена масса эректильной ткани овальной формы, называемая луковицей преддверия. Она представлена густым сплетением вен и соответствует губчатому телу полового члена у мужчин. Каждая луковица прикрепляется к нижней фасции мочеполовой диафрагмы и покрыта бульбоспонгиозной (бульбокавернозной) мышцей.
Преддверие влагалища (vestibulum vaginae) расположено между малыми половыми губами, где влагалище открывается в виде вертикальной щели. Раскрытое влагалище (так называемое отверстие) обрамляют узлы фиброзной ткани изменяющихся размеров (гименальные бугорки). Спереди от влагалищного отверстия приблизительно на 2 см ниже головки клитора по средней линии располагается наружное отверстие мочеиспускательного канала в виде небольшой вертикальной щели. Края наружного отверстия уретры обычно приподняты и образуют складки. С каждой стороны наружного отверстия мочеиспускательного канала есть миниатюрные отверстия протоков желез мочеиспускательного канала (ductus paraurethrales). Небольшое пространство в преддверии влагалища, находящееся сзади влагалищного отверстия, называется ямкой преддверия влагалища. Здесь на обеих сторонах открываются протоки больших желез преддверия, или бартолиновых желез (glandulae vestibulares majorus). Железы представляют собой небольшие дольковые тела размером с горошину и находятся у заднего края луковицы преддверия. Эти железы наряду с многочисленными малыми преддверными железами также открываются в преддверие влагалища.
Внутренние половые органы (genitalia interna). К внутренним половым органам относят влагалище, матку и ее придатки - маточные трубы и яичники (рис. 2.2).
Влагалище (vagina s. colpos) простирается от половой щели до матки, проходя кверху с задним наклонением через мочеполовую и тазовую диафрагмы (рис. 2.3). Длина влагалища около 10 см. Оно расположено главным образом в полости малого таза, где и заканчивается, сливаясь с шейкой матки. Передняя и задняя стенки влагалища обычно соединяются друг с другом в нижней части, имея форму буквы Н в поперечном сечении. Верхний отдел называют сводом влагалища, так как просвет образует карманы, или своды, вокруг влагалищной части шейки матки. Поскольку влагалище находится под углом 90° к матке, задняя стенка значительно длиннее, чем передняя, а задний свод глубже, чем передний и боковые своды. Боковая стенка влагалища прикрепляется к кардиальной связке матки и к диафрагме малого таза. Стенка состоит в основном из гладкомышечной и плотной соединительной ткани со множеством эластических волокон. Наружный слой содержит соединительную ткань с артериями, нервами и нервными сплетениями. Слизистая оболочка имеет поперечные и продольные складки. Передняя и задняя продольные складки называются столбами складок. Многослойный плоский эпителий поверхности претерпевает циклические изменения, которые соответствуют менструальному циклу.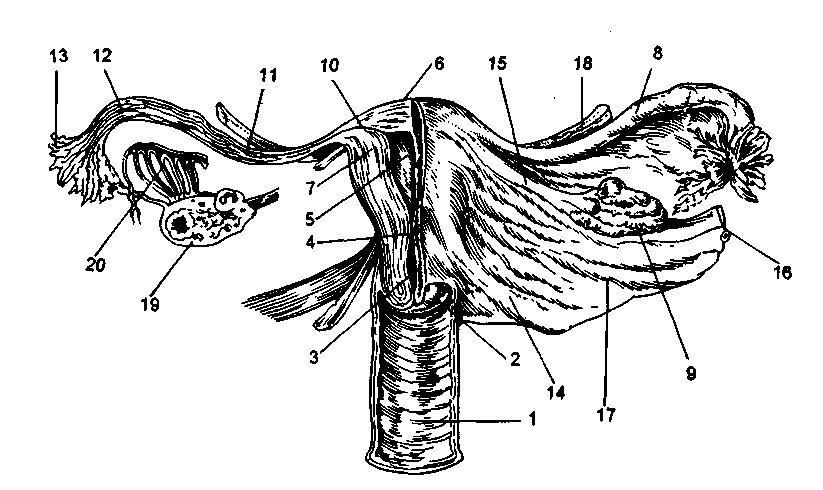
Рис. 2.2. Внутренние половые органы.
1 - влагалище; 2 - влагалищная часть шейки матки; 3 - шеечный канал; 4 - перешеек; 5 - полость матки; 6 - дно матки; 7 - стенка матки; 8 - маточная труба; 9 - яичник; 10 - интерстициальная часть трубы; 11 - истмическая часть трубы; 12 - ампулярная часть трубы; 13 - фимбрии трубы; 14 - крестцово-маточная связка; 15 - собственная связка яичника; 16 - воронкотазовая связка; 17 - широкая связка; 18 - круглая связка; 19 - срез яичника с фолликулами и желтым телом; 20 - паровариум.
Передняя стенка влагалища прилегает к мочеиспускательному каналу и основанию мочевого пузыря, причем конечная часть мочеиспускательного канала вдается в его нижнюю часть. Тонкий слой соединительной ткани, отделяющей переднюю стенку влагалища от мочевого пузыря, называется пузырно-влагалищной перегородкой. Спереди влагалище косвенно соединяется с задней частью лобковой кости с помощью фасциальных утолщений у основания мочевого пузыря, известных как лобково-пузырные связки. Сзади нижняя часть стенки влагалища отделена от заднепроходного канала перинеальным телом. Средняя часть примыкает к прямой кишке, а верхняя - к прямокишечно-маточному углублению (дугласово пространство) перитонеальной полости, от которой она отделена только тонким слоем брюшины.
Матка (uterus) вне беременности расположена по средней линии таза или вблизи нее между мочевым пузырем спереди и прямой кишкой сзади (см. рис. 2.3). Матка имеет форму перевернутой груши с плотными мышечными стенками и просветом в виде треугольника, узким в сагиттальной плоскости и широким - во фронтальной. В матке различают тело, дно, шейку и перешеек. Линия прикрепления влагалища разделяет шейку на влагалищный (вагинальный) и надвлагалищный (суправагинальный) сегменты. Вне беременности выгнутое дно направлено кпереди, причем тело образует тупой угол по отношению к влагалищу (наклонено вперед) и согнуто кпереди. Передняя поверхность тела матки плоская и примыкает к верхушке мочевого пузыря. Задняя поверхность выгнута и обращена сверху и сзади к прямой кишке.
Шейка матки направлена книзу и кзади и соприкасается с задней стенкой влагалища. Мочеточники подходят непосредственно латерально к шейке матки сравнительно близко.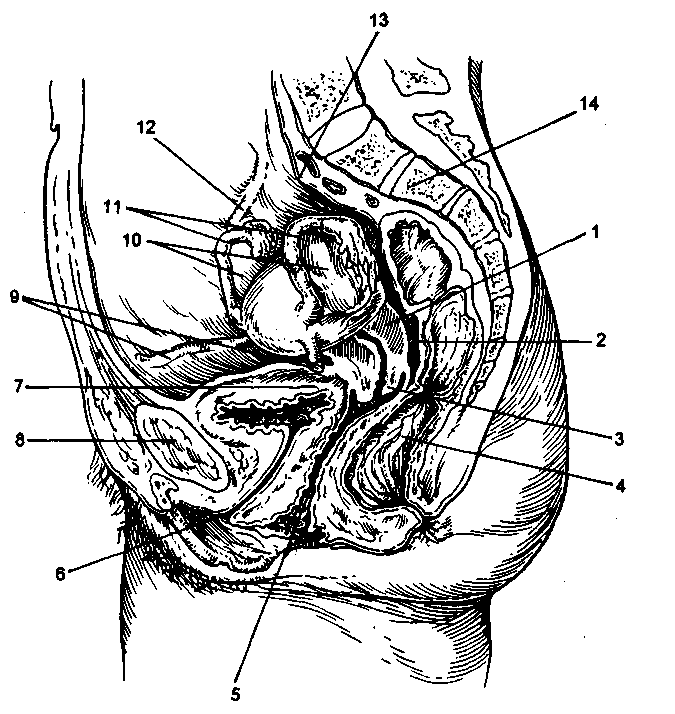
Рис. 2.3. Соотношение органов малого таза (сагиттальный срез).
1 - матка; 2 - прямокишечно-маточное углубление; 3 - шейка матки; 4 - прямая кишка; 5 - влагалище; 6 - мочеиспускательный канал; 7 - мочевой пузырь; 8 - симфиз; 9 - круглые связки матки; 10 - яичники; И -маточные трубы; 12 - воронкотазовая связка; 13 - крестцовый мыс; 14 - крестец.
Тело матки, включая ее дно, покрыто брюшиной. Спереди, на уровне перешейка, брюшина загибается и переходит на верхнюю поверхность мочевого пузыря, образуя неглубокое пузырно-маточное углубление. Сзади брюшина продолжается вперед и вверх, покрывая перешеек, суправагинальную часть шейки матки и задний свод влагалища, а затем переходит на переднюю поверхность прямой кишки, образуя глубокое прямокишечно-маточное углубление. Длина тела матки в среднем равна 5 см. Общая длина перешейка и шейки около 2,5 см, их диаметр 2 см. Соотношение длины тела и шейки матки зависит от возраста и числа родов и в среднем составляет 2:1.
Стенка матки состоит из тонкого наружного слоя брюшины - серозной оболочки (периметрии), толстого промежуточного слоя гладких мышц и соединительной ткани - мышечной оболочки (миометрий) и внутренней слизистой оболочки (эндометрий). Тело матки содержит множество мышечных волокон, количество которых уменьшается книзу по мере приближения к шейке. Шейка состоит из равного количества мышц и соединительной ткани. В результате своего развития из слившихся частей парамезонефральных (мюллеровых) протоков расположение мышечных волокон в стенке матки сложное. Наружный слой миометрия содержит в основном вертикальные волокна, которые идут латерально в верхней части тела и соединяются с наружным продольным мышечным слоем маточных труб. Средний слой включает в себя большую часть маточной стенки и состоит из сети спиралевидных мышечных волокон, которые соединены с внутренним круговым мышечным слоем каждой трубы. Пучки гладкомышечных волокон в поддерживающих связках переплетаются и сливаются с этим слоем. Внутренний слой состоит из круговых волокон, которые могут выполнять функцию сфинктера у перешейка и у отверстий маточных труб.
Полость матки вне беременности представляет собой узкую щель, при этом передняя и задняя стенки тесно прилегают друг к другу. Полость имеет форму перевернутого треугольника, основание которого находится сверху, где оно с обеих сторон соединено с отверстиями маточных труб; вершина находится снизу, где полость матки переходит в шеечный канал. Шеечный канал в области перешейка сжат и имеет длину 6-10 мм. Место, где канал шейки матки переходит в полость матки, называется внутренним зевом. Цервикальный канал немного расширяется в своей средней части и открывается во влагалище наружным отверстием.
Придатки матки. К придаткам матки относят маточные трубы и яичники, а некоторые авторы - и связочный аппарат матки.
Маточные трубы (tubae uterinae). С обеих сторон тела матки латерально находятся длинные, узкие маточные трубы (фаллопиевы трубы). Трубы занимают верхнюю часть широкой связки и изгибаются дугой латерально над яичником, затем идут вниз над задней частью медиальной поверхности яичника. Просвет, или канал, трубы проходит от верхнего угла полости матки к яичнику, постепенно увеличиваясь в диаметре латерально по его ходу. Вне беременности труба в растянутом виде имеет длину 10 см. Различают четыре ее отдела: интрамуральный участок находится внутри стенки матки и соединен с полостью матки. Его просвет имеет самый маленький диаметр (1 мм или менее). Узкий участок, идущий латерально от наружной границы матки, называется перешейком (istmus); далее труба расширяется и становится извилистой, образуя ампулу, и заканчивается вблизи яичника в виде воронки. По периферии на воронке находятся фимбрии, которые окружают брюшное отверстие маточной трубы; одна или две фимбрии соприкасаются с яичником. Стенка маточной трубы образована тремя слоями: наружный слой, состоящий в основном из брюшины (серозной оболочки), промежуточный гладкий мышечный слой (миосальпинкс) и слизистая оболочка (эндосальпинкс). Слизистая оболочка представлена реснитчатым эпителием и имеет продольные складки.
Яичники (ovarii). Женские гонады представлены яичниками овальной или миндалевидной формы. Яичники расположены медиально к загнутой части маточной трубы и немного расплющены. В среднем их размеры составляют: ширина 2 см, длина 4 см и толщина 1 см. Яичники, как правило, серовато-розового цвета с морщинистой, неровной поверхностью. Продольная ось яичников почти вертикальная, с верхней крайней точкой у маточной трубы и с нижней крайней точкой ближе к матке. Задняя часть яичников свободна, а передняя фиксирована к широкой связке матки с помощью двухслойной складки брюшины - брыжейки яичника (mesovarium). Через нее проходят сосуды и нервы, которые достигают ворот яичников. К верхнему полюсу яичников прикреплены складки брюшины - связки, подвешивающие яичники (воронкотазовые), в которых содержатся яичниковые сосуды и нервы. Нижняя часть яичников прикреплена к матке с помощью фиброзно-мышечных связок (собственные связки яичников). Эти связки соединяются с латеральными краями матки под углом чуть ниже того места, где маточная труба подходит к телу матки.
Яичники покрыты зародышевым эпителием, под которым находится слой соединительной ткани - белочная оболочка. В яичнике различают наружный корковый и внутренний мозговой слои. В соединительной ткани мозгового слоя проходят сосуды, нервы. В корковом слое среди соединительной ткани находится большое количество фолликулов на разных стадиях развития.
Связочный аппарат внутренних женских половых органов. Положение в малом тазе матки и яичников, а также влагалища и смежных органов зависит главным образом от состояния мышц и фасций тазового дна, а также от состояния связочного аппарата матки (см. рис. 2.2). В нормальном положении матку с маточными трубами и яичники удерживают подвешивающий аппарат (связки), закрепляющий аппарат (связки, фиксирующие подвешенную матку), опорный, или поддерживающий, аппарат (тазовое дно).
Подвешивающий аппарат внутренних половых органов включает следующие связки.
1. Круглые связки матки (ligg. teres uteri). Они состоят из гладких мышц и соединительной ткани, имеют вид шнуров длиной 10-12 см. Эти связки отходят от углов матки, идут под передним листком широкой связки матки к внутренним отверстиям паховых каналов. Пройдя паховый канал, круглые связки матки веерообразно разветвляются в клетчатке лобка и больших половых губ. Круглые связки матки притягивают дно матки кпереди (наклонение кпереди).
2. Широкие связки матки (ligg. latae uteri). Это дупликатура брюшины, идущая от ребер матки до боковых стенок таза. В верхних отделах широких
Связок матки проходят маточные трубы, на задних листках расположены яичники, между листками - клетчатка, сосуды и нервы.
3. Собственные связки яичников (ligg. ovarii proprii, s. ligg. suspensorii ovarii) начинаются от дна матки сзади и ниже места отхождения маточных труб и идут к яичникам.
4. Связки, подвешивающие яичники, или воронкотазовые связки (ligg. suspensorium ovarii, s.infundibulopelvicum), являются продолжением широких маточных связок, идут от маточной трубы до стенки таза.
Закрепляющий аппарат матки представляет собой соединительнотканные тяжи с примесью гладкомышечных волокон, которые идут от нижнего отдела матки:
Б) кзади - к прямой кишке и крестцу (lig. sacrouterinum).
Крестцово-маточные связки отходят от задней поверхности матки в области перехода тела в шейку, охватывают с обеих сторон прямую кишку и прикрепляются на передней поверхности крестца. Эти связки притягивают шейку матки кзади.
Опорный, или поддерживающий, аппарат составляют мышцы и фасции тазового дна. Тазовое дно имеет огромное значение в удержании внутренних половых органов в нормальном положении. При повышении внутрибрюшного давления шейка матки опирается на тазовое дно, как на подставку; мышцы тазового дна препятствуют опусканию вниз половых органов и внутренностей. Тазовое дно образовано кожей и слизистой оболочкой промежности, а также мышечно-фасциальной диафрагмой.
Промежность (perineum) - это ромбовидная область между бедрами и ягодицами, где находятся мочеиспускательный канал, влагалище и заднепроходное отверстие. Спереди промежность ограничена лобковым симфизом, сзади - концом копчика, латерально-седалищными буграми. Кожа ограничивает промежность снаружи и снизу, а диафрагма таза (тазовая фасция), образованная нижней и верхней фасциями, ограничивает промежность глубоко сверху (рис. 2.4).
Тазовое дно с помощью воображаемой линии, соединяющей два седалищных бугра, разделяют анатомически на две треугольные области: спереди - мочеполовая область, сзади - анальная область. В центре промежности между заднепроходным отверстием и входом во влагалище имеется фиброзно-мышечное образование, называемое сухожильным центром промежности. Этот сухожильный центр является местом прикрепления нескольких групп мышц и фасциальных слоев.
Мочеполовая область. В мочеполовой области между нижними ветвями седалищных и лобковых костей находится мышечно-фасциальное образование, которое называется "мочеполовая диафрагма" (diaphragma urogenitale). Через эту диафрагму проходят влагалище и мочеиспускательный канал. Диафрагма служит основанием для фиксации наружных половых органов. Снизу мочеполовая диафрагма ограничена поверхностью беловатых коллагеновых волокон, образующих нижнюю фасцию мочеполовой диафрагмы, которая делит мочеполовую область на два плотных анатомических слоя, имеющих важное клиническое значение, - поверхностный и глубокий отделы, или карманы промежности.
Поверхностный отдел промежности.
Поверхностный отдел находится над нижней фасцией мочеполовой диафрагмы и содержит на каждой стороне большую железу преддверия влагалища, ножку клитора с лежащей сверху седалищно-пещеристой мышцей, луковицу преддверия с лежащей сверху луковично-губчатой (луковично-пещеристой) мышцей и небольшую поверхностную поперечную мышцу промежности. Седалищно-пещеристая мышца покрывает ножку клитора и играет значительную роль в сохранении его эрекции, так как прижимает ножку к седалищно-лобковой ветви, задерживая отток крови из эректальной ткани. Луковично-губчатая мышца начинается от сухожильного-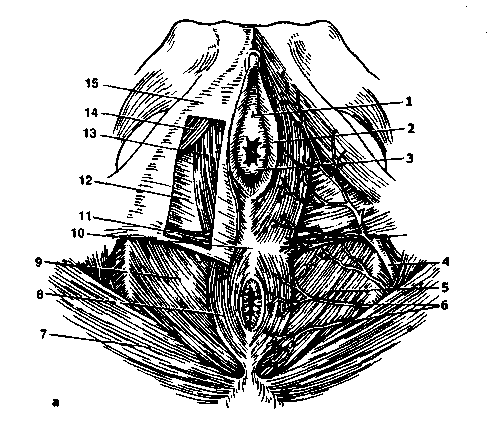
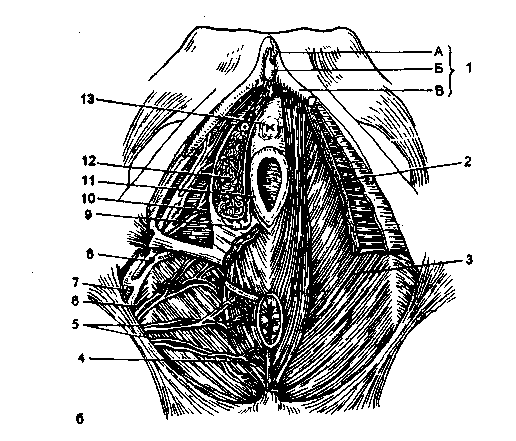
Рис. 2.4. Женская промежность.
А - поверхностный срез мочеполовой диафрагмы: 1 - наружное отверстие мочеиспускательного канала, 2 - малая половая губа, 3 - девственная плева, 4 - внутренняя половая артерия, 5 - мышцы, поднимающие задний проход, 6 - нижняя геморроидальная артерия, 7 - большая ягодичная мышца, 8 - наружный сфинктер заднего прохода, 9 - нижняя фасция диафрагмы таза, 10 - сухожильный центр промежности, 11 -наружная поперечная мышца промежности, 12 - нижняя фасция мочеполовой диафрагмы, 13 - луковично-губчатая мышца, 14 - седалищно-пещеристая мышца, 15 - поверхностная фасция промежности; б*- глубокий срез мочеполовой диафрагмы: 1 - клитор: А - тело, Б - головка, В - ножка; 2 - урогенитальная диафрагма, 3 - тазовая диафрагма, 4 - мышца наружного сфинктера заднего прохода, 5 - нижняя геморроидальная артерия, 6 - внутренняя запирательная мышца, 7 - внутренняя половая артерия, 8 - артерия промежности, 9 - большая железа преддверия, 10 - преддвериелуковичная артерия, 11 - стенка влагалища, 12- луковица преддверия, 13 - мочеиспускательный канал.
центра промежности и наружного сфинктера заднего прохода, затем проходит сзади вокруг нижней части влагалища, покрывая луковицу преддверия, и входит в перинеальное тело. Мышца может действовать в качестве сфинктера для сжатия нижней части влагалища. Слабо развитая, имеющая вид тонкой пластинки поверхностная поперечная мышца промежности начинается от внутренней поверхности седалищной кости около седалищного бугра и идет поперечно, входя в перинеальное тело. Все мышцы поверхностного отдела покрыты глубокой фасцией промежности.
Глубокий отдел промежности. Глубокий отдел промежности находится между нижней фасцией мочеполовой диафрагмы и неотчетливо выраженной верхней фасцией мочеполовой диафрагмы. Мочеполовая диафрагма состоит из двух слоев мышц. Мышечные волокна в мочеполовой диафрагме в основном располагаются поперечно, отходят от седалищно-лобковых ветвей каждой стороны и соединяются по средней линии. Эта часть мочеполовой диафрагмы называется глубокой поперечной мышцей промежности (m. transversus perinei profundus). Часть волокон сфинктера мочеиспускательного канала поднимается дугой над мочеиспускательным каналом, тогда как другая часть расположена вокруг него циркулярно, образуя наружный сфинктер уретры. Мышечные волокна сфинктера мочеиспускательного канала также проходят вокруг влагалища, концентрируясь там, где располагается наружное отверстие мочеиспускательного канала. Мышца играет важную роль в сдерживании процесса мочеиспускания при наполненном мочевом пузыре и является произвольным сжимателем мочеиспускательного канала. Глубокая поперечная мышца промежности входит в перинеальное тело позади влагалища. При билатеральном сокращении эта мышца поддерживает, таким образом, промежность и висцеральные структуры, проходящие через нее.
Вдоль переднего края мочеполовой диафрагмы сливаются две ее фасции, образуя поперечную связку промежности. Спереди этого фасциального утолщения находится дугообразная лобковая связка, проходящая вдоль нижнего края лобкового симфиза.
Заднепроходная (анальная) область. Заднепроходная (анальная) область включает в себя задний проход (anus), наружный сфинктер заднего прохода и седалищно-прямокишечную ямку. Задний проход расположен на поверхности промежности. Кожа заднего прохода пигментирована и содержит сальные и потовые железы. Сфинктер заднего прохода состоит из поверхностной и глубокой частей поперечно-полосатых мышечных волокон. Подкожная часть является самой поверхностной и окружает нижнюю стенку прямой кишки, глубокая часть состоит из круговых волокон, которые сливаются с мышцей, поднимающей задний проход (m.levator ani). Поверхностная часть сфинктера состоит из мышечных волокон, идущих в основном вдоль анального канала и пересекающихся под прямым углом впереди и позади заднепроходного отверстия, которые затем попадают спереди в промежность, а сзади - в слабо выраженную фиброзную массу, называемую анально-копчиковым телом, или анально-копчиковой связкой. Задний проход внешне представляет собой продольное щелевидное отверстие, что объясняется, возможно, переднезадним направлением многих мышечных волокон наружного сфинктера заднего прохода.
Седалигцно-прямокишечная ямка представляет собой клиновидное пространство, наполненное жиром, которое снаружи ограничено кожей. Кожа образует основу клина. Вертикальная боковая стенка ямки образована внутренней запирательной мышцей. Наклонная супрамедиальная стенка содержит мышцу, поднимающую задний проход. Седалищно-прямокишечная жировая клетчатка позволяет прямой кишке и анальному каналу расширяться во время дефекации. Ямка и содержащаяся в ней жировая клетчатка располагаются кпереди и глубоко кверху до мочеполовой диафрагмы, но ниже мышцы, поднимающей задний проход. Эта зона называется передним карманом. Сзади жировая клетчатка в ямке проходит глубоко до большой ягодичной мышцы в зоне крестцово-бугорной связки. Латерально ямка ограничена седалищной костью и запирательной фасцией, покрывающей нижнюю часть внутренней запирательной мышцы.
Кровоснабжение, лимфоотток и иннервация половых органов. Кровоснабжение (рис. 2.5, 2.6) наружных половых органов в основном осуществляется внутренней половой (срамной) артерией и лишь частично веточками бедренной артерии.
Внутренняя половая артерия (a.pudenda interna) является главной артерией промежности. Она представляет собой одну из ветвей внутренней подвздошной артерии (a.iliaca interna). Покидая полость малого таза, она проходит в нижней части большого седалищного отверстия, затем огибает седалищную ость и идет по боковой стенке седалишно-ректальной ямки, поперечно пересекая малое седалищное отверстие. Первой ее ветвью является нижняя прямокишечная артерия (a.rectalis inferior). Проходя через седалищно-ректальную ямку, она снабжает кровью кожу и мышцы вокруг заднего прохода. Промежностная ветвь снабжает структуры поверхностного отдела промежности и продолжается в виде задних ветвей, идущих к большим и малым половым губам. Внутренняя половая артерия, входя в глубокий промежностный отдел, разветвляется на несколько фрагментов и кровоснабжает луковицу преддверия влагалища, большую железу преддверия и мочеиспускательного канала. Заканчиваясь, она разделяется на глубокую и дорсальную артерии клитора, подходящие к нему около лобкового симфиза.
Наружная (поверхностная) половая артерия (r.pudenda externa, s.superficialis) отходит от медиальной стороны бедренной артерии (a.femoralis) и кровоснабжает переднюю часть больших половых губ. Наружная (глубокая) половая артерия (r.pudenda externa, s.profunda) также отходит от бедренной артерии, но более глубоко и дистальнее. Пройдя широкую фасцию на медиальной стороне бедра, она входит в латеральную часть большой половой губы. Ее ветви переходят в передние и задние лабиальные артерии.
Вены, проходящие через промежность, являются в основном ветвями внутренней подвздошной вены. Большей частью они сопровождают артерии. Исключение составляет глубокая дорсальная вена клитора, которая отводит кровь из эректильной ткани клитора через щель ниже лобкового симфиза в венозное сплетение вокруг шейки мочевого пузыря. Наружные половые вены отводят кровь от большой половой губы, проходя латерально и входя в большую подкожную вену ноги.
Кровоснабжение внутренних половых органов осуществляется в основном из аорты (система общей и внутренней подвздошной артерий).
Основное кровоснабжение матки обеспечивается маточной артерией (a.uterina), которая отходит от внутренней подвздошной (подчревной) артерии (a.iliaca interna). Примерно в половине случаев маточная артерия самостоятельно отходит от внутренней подвздошной артерии, но она может начинаться и от пупочной, внутренней половой и поверхностной пузырной артерий.
Маточная артерия направляется вниз к боковой тазовой стенке, затем проходит вперед и медиально, располагаясь над мочеточником, к которому может давать самостоятельную ветвь. У основания широкой маточной связки она поворачивает медиально по направлению к шейке матки. В параметрии артерия соединяется с сопровождающими ее венами, нервами, мочеточником и кардинальной связкой. Маточная артерия подходит к шейке матки и снабжает ее с помощью нескольких извилистых проникающих ветвей. Затем маточная артерия разделяется на одну большую очень изви-листую восходящую ветвь и одну или несколько мелких нисходящих ветвей, снабжающих верхнюю часть влагалища и прилегающую часть мочевого пузыря. Главная восходящая ветвь идет вверх вдоль латерального края матки, посылая дугообразные ветви к ее телу.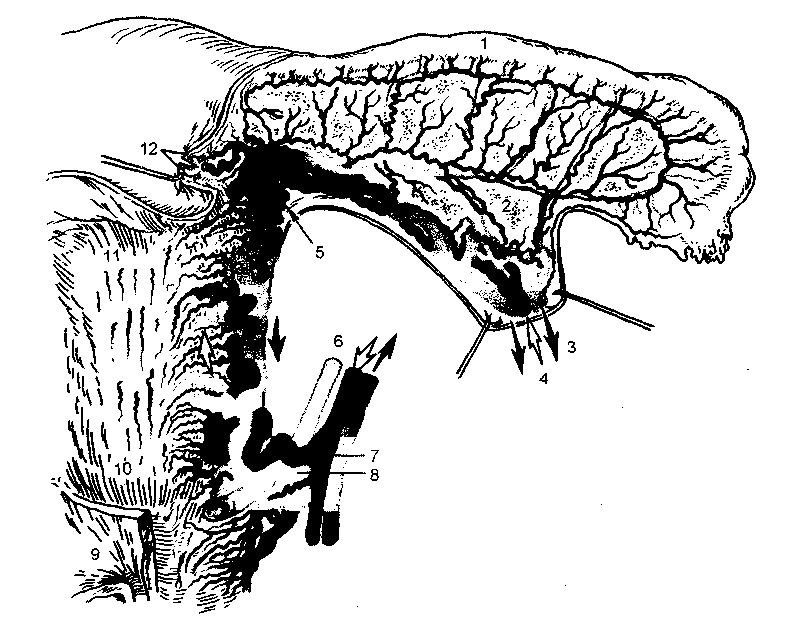
Рис. 2.5. Кровоснабжение половых органов.
1 - маточная труба; 2 - яичник; 3 - яичниковая вена; 4 - яичниковая артерия; 5 - анастомозы маточных и яичниковых сосудов; 6 - мочеточник; 7 - маточная артерия; 8 - маточная вена; 9 - стенка мочевого пузыря; 10 - шейка матки; 11 - тело матки; 12 - круглая связка матки.
Эти дугообразные артерии окружают матку под серозным слоем. С определенными промежутками от них отходят радиальные ветви, которые проникают в переплетающиеся мышечные волокна миометрия. После родов мышечные волокна сокращаются и, действуя как лигатуры, сжимают радиальные ветви. Дугообразные артерии быстро уменьшаются в размере по ходу к средней линии, поэтому при срединных разрезах матки наблюдается меньшее кровотечение, чем при латеральных. Восходящая ветвь маточной артерии подходит к маточной трубе, поворачивая латерально в ее верхней части, и разделяется на трубную и яичниковую ветви. Трубная ветвь идет латерально в брыжейке маточной трубы (mesosalpinx). Яичниковая ветвь направляется к брыжейке яичника (mesovarium), где она анастомозирует с яичниковой артерией, отходящей непосредственно от аорты.
Яичники кровоснабжаются из яичниковой артерии (a.ovarica), отходящей от брюшной аорты слева, иногда от почечной артерии (a.renalis). Спускаясь вместе с мочеточником вниз, яичниковая артерия проходит по связке, подвешивающей яичник, к верхнему отделу широкой маточной связки, отдает ветвь для яичника и трубы; конечный отдел яичниковой артерии анастомозирует с концевым отделом маточной артерии.
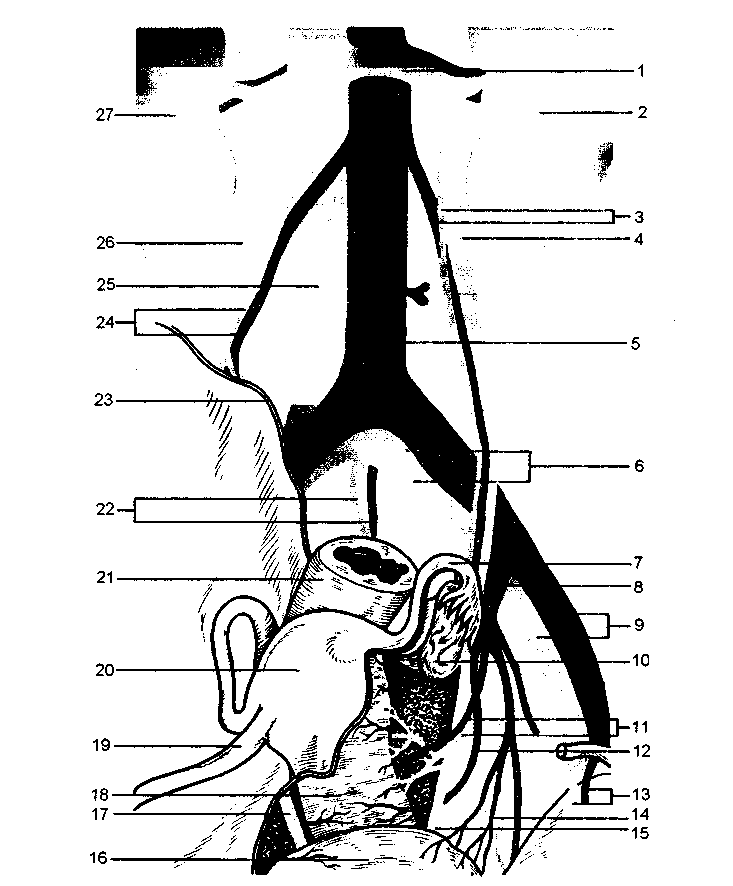
Рис. 2.6. Взаимоотношение внутренних половых органов, мочеточников и сосудов малого таза.
1 - левая почечная вена; 2 - левая почка; 3 - левые яичниковые вена и артерия; 4 - левый мочеточник; 5 - брюшная часть аорты; 6 - общие подвздошные артерия и вена; 7 - маточная труба; 8 - внутренняя подвздошная артерия; 9 - наружные подвздошные артерия и вена; 10 - левый яичник; 11 - маточные артерия и вена; 12 - нижняя пузырная артерия (влагалищная ветвь); 13 - нижние надчревные артерия и вена; 14 -верхняя мочепузырная артерия; 15 -левый мочеточник; 16 - мочевой пузырь; 17 - правый мочеточник; 18 - влагалище; 19 - круглая связка матки; 20 - тело матки; 21 - прямая кишка; 22 - средние крестцовые вена и артерия; 23 - край париетальной брюшины (в разрезе); 24 - правые яичниковые артерия и вена; 25 - нижняя полая вена; 26 - правый мочеточник; 27 - правая почка.
В кровоснабжении влагалища, кроме маточной и половой артерий, участвуют также ветви нижней мочепузырной и средней прямокишечной артерии. Артерии половых органов сопровождаются соответствующими венами.
Венозная система "половых органов развита очень сильно; общая протяженность венозных сосудов... значительно превышает протяженность артерий в связи с наличием веноз.ных сплетений, широко анастомозирующих между собой. Венозные сплетения находятся в области клитора, у краев луковиц преддверия, вокруг мочевого пузыря, между маткой и яичниками.
Лимфатическая система половых органов состоит из густой сети извилистых лимфатических сосудов, сплетений и множества лимфатических узлов. Лимфатические пути и узлы располагаются преимущественно по ходу кровеносных сосудов.
Лимфатические сосуды, отводящие лимфу от наружных половых органов и нижней трети влагалища, идут к паховым лимфатическим узлам. Лимфатические пути, отходящие от средней верхней трети влагалища и шейки матки, идут к лимфатическим узлам, располагающимся по ходу подчревных и подвздошных кровеносных сосудов.
Интрамуральные сплетения несут лимфу из эндометрия и миометрия в субсерозное сплетение, из которого лимфа оттекает по эфферентным сосудам. Лимфа из нижней части матки поступает в основном в крестцовые, наружные подвздошные и общие подвздошные лимфатические узлы; часть лимфы также поступает в нижние поясничные узлы вдоль брюшной аорты и в поверхностные паховые узлы. Большая часть лимфы из верхней части матки оттекает в латеральном направлении в широкой связке матки, где она соединяется с лимфой, собирающейся из маточной трубы и яичника. Далее через связку, подвешивающую яичник, по ходу яичниковых сосудов лимфа поступает в лимфатические узлы вдоль нижнего отдела брюшной аорты. Из яичников лимфа отводится по сосудам, располагающимся вдоль яичниковой артерии, и идет к лимфатическим узлам, лежащим на аорте и нижней полой вене. Между указанными лимфатическими сплетениями существуют связи - лимфатические анастомозы.
В иннервации половых органов женщины участвуют симпатическая и парасимпатическая части вегетативной нервной системы, а также спинномозговые нервы.
Волокна симпатической части вегетативной нервной системы, иннервирующие половые органы, берут начало от аортального и чревного ("солнечного") сплетений, направляются вниз и на уровне V поясничного позвонка образуют верхнее подчревное сплетение (plexus hypogastricus superior). От него отходят волокна, образующие правое и левое нижние подчревные сплетения (plexus hypogastricus sinister et dexter inferior). Нервные волокна от этих сплетений идут к мощному маточно-влагалищному, или тазовому, сплетению (plexus uterovaginalis, s.pelvicus).
Маточно-влагалищные сплетения располагаются в параметральной клетчатке сбоку и сзади от матки на уровне внутреннего зева и шеечного канала. К этому сплетению подходят ветви тазового нерва (n.pelvicus), относящегося к парасимпатической части вегетативной нервной системы. Симпатические и парасимпатические волокна, отходящие от маточно-влагалищного сплетения, иннервируют влагалище, матку, внутренние отделы маточных труб, мочевой пузырь.
Яичники иннервируются симпатическими и парасимпатическими нервами из яичникового сплетения (plexus ovaricus).
Наружные половые органы и тазовое дно в основном иннервируются половым нервом (n.pudendus).
Тазовая клетчатка. Кровеносные сосуды, нервы и лимфатические пути органов малого таза проходят в клетчатке, которая расположена между брюшиной и фасциями тазового дна. Клетчатка окружает все органы малого таза; на одних участках она рыхлая, на других в виде волокнистых тяжей. Различают следующие пространства клетчатки: околоматочное, пред- и околопузырное, околокишечное, влагалищное. Тазовая клетчатка служит опорой для внутренних половых органов, и все ее отделы связаны между собой.
2.1.3. Таз с акушерской точки зрения
Большой таз для рождения ребенка существенного значения не имеет. Костную основу родового канала, представляющего препятствие рождающемуся плоду, составляет малый таз. Однако по размерам большого таза можно косвенно судить о форме и величине малого таза. Внутреннюю поверхность большого и малого таза выстилают мышцы.
Полостью малого таза
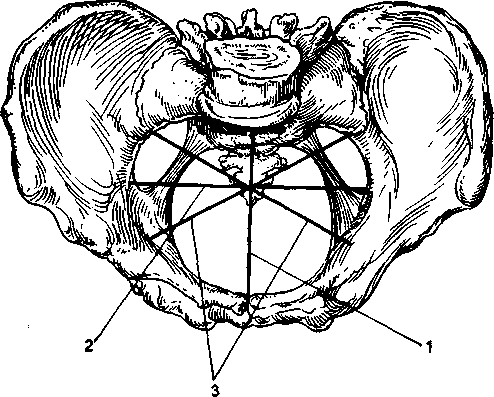
называется пространство, заключенное между стенками таза, сверху и снизу ограниченное плоскостями входа и выхода таза. Она имеет вид цилиндра, усеченного спереди назад, причем передняя часть, обращенная к лону, почти в 3 раза ниже задней, обращенной к крестцу. В связи с такой формой полости малого таза различные ее отделы имеют неодинаковую форму и размеры. Этими отделами являются воображаемые плоскости, проходящие через опознавательные пункты внутренней поверхности малого таза. В малом тазе различают следующие плоскости: плоскость входа, плоскость широкой части, плоскость узкой части и плоскость выхода (табл. 2.1; рис. 2.7).
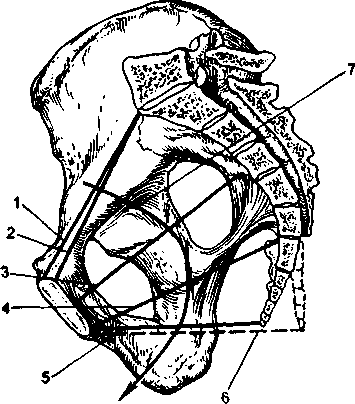 |  |
| Рис. 2.7. Женский таз (сагиттальный срез). 1 - анатомическая конъюгата; 2 - истинная конъюгата; 3 - прямой размер плоскости широкой части полости малого таза; 4 - прямой размер плоскости узкой части полости малого таза; 5 - прямой размер выхода малого таза при обычном положени копчика; 6 - прямой размер выхода малого таза при отогнутом кзади копчике; 7 - проводная ось таза. | Рис. 2.8. Размеры плоскости входа в малый таз. 1 - прямой размер (истинная конъюгата); 2 - поперечный размер; 3 - косые размеры. |
Плоскость входа в малый таз проходит через верхневнутренний край лонной дуги, безымянные линии и вершину мыса. В плоскости входа различают следующие размеры (рис. 2.8).
Прямой размер- кратчайшее расстояние между серединой верхневнутреннего края лонной дуги и самой выдающейся точкой мыса. Это расстояние называется истинной конъюгатой (conjugata vera); она равняется 11 см. Принято также различать и анатомическую конъюгату - расстояние от середины верхнего края лонной дуги до той же точки мыса; она на 0,2-0,3 см длиннее истинной конъюгаты (см. рис. 2.7).
Поперечный размер - расстояние между наиболее отдаленными точками безымянных линий противоположных сторон. Он равен 13,5 см. Этот размер пересекает под прямым углом истинную конъюгату эксцентрично, ближе к мысу.
Косые размеры- правый и левый. Правый косой размер идет от правого крестцово-подвздошного сочленения до левого подвздошно-лонного бугорка, а левый косой размер - соответственно от левого крестцово-подвздошного сочленения до правого подвздошно-лонного бугорка. Каждый из этих размеров равен 12 см.
Как видно из приведенных размеров, плоскость входа имеет поперечно-овальную форму.
Плоскость широкой части полости малого таза проходит спереди через середину внутренней поверхности лонной дуги, с боков - через середину гладких пластинок, расположенных под ямками вертлужных впадин (lamina acetabuli), и сзади - через сочленение между II и III крестцовыми позвонками.
В плоскости широкой части различают следующие размеры.
Прямой размер- от середины внутренней поверхности лонной дуги до сочленения между II и III крестцовыми позвонками; он равен 12,5 см.
Поперечный размер, соединяющий наиболее отдаленные точки пластинок вертлужных впадин той и другой стороны равен 12,5 см.
Плоскость широкой части по своей форме приближается к кругу.
Плоскость узкой части полости малого таза проходит спереди через нижний край лонного сочленения, с боков - через седалищные ости, сзади - через крестцово-копчиковое сочленение.
В плоскости узкой части различают следующие размеры.
Прямой размер- от нижнего края лонного сочленения к крестцово-копчиковому сочленению. Он равен 11 см.
Поперечный размер -между внутренней поверхностью седалищных остей. Он равен 10,5 см.
Плоскость выхода малого таза в отличие от других плоскостей малого таза состоит из двух плоскостей, сходящихся под углом по линии, соединяющей седалищные бугры. Она проходит спереди через нижний край лонной дуги, по бокам - через внутренние поверхности седалищных бугров и сзади - через верхушку копчика.
В плоскости выхода различают следующие размеры.
Прямой размер- от середины нижнего края лонного сочленения до верхушки копчика. Он равен 9,5 см (рис. 2.9). Прямой размер выхода благодаря некоторой подвижности копчика может удлиняться в родах при прохождении головки плода на 1-2 см и достигать 11,5 см (см. рис. 2.7).
Поперечный размер -между наиболее отдаленными точками внутренних поверхностей седалищных бугров. Он равен 11 см (рис. 2.10).
Таблица 2.1. Плоскости и размеры малого таза
| Плоскости таза | Размеры, см |
|||||
| прямой | поперечный | косой |
||||
| Вход в таз | 11 | 13-13,5 | 12-12,5 |
|||
| Широкая часть полости таза | 12,5 | 12,5 | 13 (условно) |
|||
| Узкая часть полости таза | 11-11,5 | 10,5 | - |
|||
| Выход таза | 9,5-11,5 | 11 | - |
|||
|
|
| |||||
| Рис. 2.9. Прямой размер выхода таза (измерение). | Рис. 2.10. Измерение поперечного размера выхода таза. | |||||
Эта классическая система плоскостей, в разработке которой приняли участие основоположники русского акушерства, особенно А.Я.Крассовский, позволяет правильно ориентироваться в продвижении предлежащей части плода по родовому каналу и совершаемых ею при этом поворотах.
В 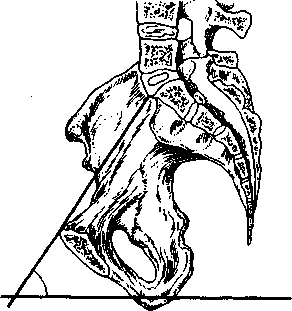
Рис. 2.11. Угол наклонения таза
се прямые размеры плоскостей малого таза сходятся в области лонного сочленения, в области же крестца расходятся. Линия, соединяющая середины всех прямых размеров плоскостей малого таза, представляет собой дугу, вогнутую спереди и выгнутую сзади. Эта линия называется проводной осью малого таза. Прохождение плода по родовому каналу совершается по этой линии (см. рис. 2.7).
Угол наклонения таза - пересечение плоскости его входа с плоскостью горизонта (рис. 2.11) - при положении женщины стоя может быть различным в зависимости от телосложения и колеблется от 45 до 55°. Он может быть уменьшен, если попросить женщину, лежащую на спине, сильно притянуть к животу бедра, что приводит к приподниманию лона, или, наоборот, увеличен, если подложить под поясницу валикообразную жесткую подушку, что приведет к отклонению лона вниз. Уменьшение угла наклонения таза достигается также в том случае, если женщина принимает положение полусидя или положение на корточ
2.2. ФИЗИОЛОГИЯ ЖЕНСКОЙ РЕПРОДУКТИВНОЙ СИСТЕМЫ. МЕНСТРУАЛЬНЫЙ ЦИКЛ
Репродуктивная функция женщин осуществляется прежде всего благодаря деятельности яичников и матки, так как в яичниках созревает яйцеклетка, а в матке под влиянием гормонов, выделяемых яичниками, происходят изменения по подготовке к восприятию оплодотворенного плодного яйца. Репродуктивный период характеризуется способностью организма женщины к воспроизводству потомства; продолжительность данного периода от 17-18 до 45-50 лет. Репродуктивному, или детородному, периоду предшествуют следующие этапы жизни женщины: внутриутробный; новорожденности (до 1 года); детства (до 8-10 лет); препубертатного и пубертатного возраста (до 17-18 лет). Репродуктивный период переходит в климактерический, в котором различают пременопаузу, менопаузу и постменопаузу.
Менструальный цикл - одно из проявлений сложных биологических процессов в организме женщины. Менструальный цикл характеризуется циклическими изменениями во всех звеньях репродуктивной системы, внешним проявлением которых является менструация.
Менструации - это кровянистые выделения из половых путей женщины, периодически возникающие в результате отторжения функционального слоя эндометрия в конце двухфазного менструального цикла. Первая менструация (menarhe) наблюдается в возрасте 10-12 лет, но в течение 1 - 1,5 года после этого менструации могут быть нерегулярными, а затем устанавливается регулярный менструальный цикл.
Первый день менструации условно принимается за первый день менструального цикла. Следовательно, продолжительность цикла составляет время между первыми днями двух последующих менструаций. Для 60 % женщин средняя продолжительность менструального цикла составляет 28 дней с колебаниями от 21 до 35 дней. Величина кровопотери в менструальные дни 40-60 мл, в среднем 50 мл. Продолжительность нормальной менструации от 2 до 7 дней.
Яичники. Во время менструального цикла в яичниках происходит рост фолликулов и созревание яйцеклетки, которая в результате становится готовой к оплодотворению. Одновременно в яичниках вырабатываются половые гормоны, обеспечивающие изменения в слизистой оболочке матки, способной воспринять оплодотворенное яйцо.
Половые гормоны (эстрогены, прогестерон, андрогены) являются стероидами, в их образовании принимают участие гранулезные клетки фолликула, клетки внутреннего и наружного слоев. Половые гормоны, синтезируемые яичниками, влияют на ткани- и органы-мишени. К ним относятся половые органы, в первую очередь матка, молочные железы, губчатое вещество костей, мозг, эндотелий и гладкие мышечные клетки сосудов, миокард, кожа и ее придатки (волосяные фолликулы и сальные железы) и др. Прямой контакт и специфическое связывание гормонов на клетке-мишени является результатом взаимодействия его с соответствующими рецепторами.
Биологический эффект дают свободные (несвязанные) фракции эстрадиола и тестостерона (1 %). Основная масса яичниковых гормонов (99 %) находится в связанном состоянии. Транспорт осуществляется специальными белками - стероидосвязывающими глобулинами и неспецифическими транспортными системами - альбуминами и эритроцитами.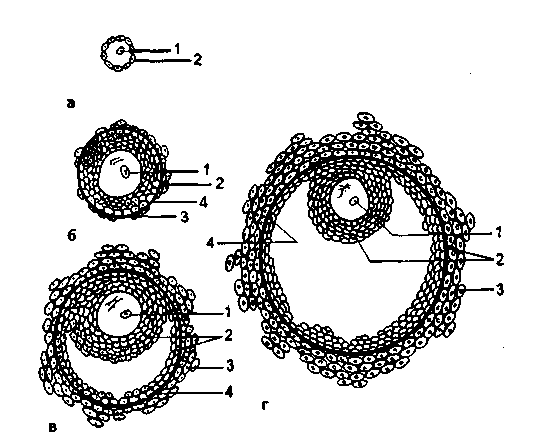
Рис. 2.12. Этапы развития доминантного фолликула.
А - примордиальный фолликул; б - преантральный фолликул; в - антральный фолликул; г - преовуляторный фолликул: 1 - овоцит, 2 - гранулезные клетки (зернистая зона), 3 - тека-клетки, 4 - базальная мембрана.
Эстрогенные гормоны способствуют формированию половых органов, развитию вторичных половых признаков в период полового созревания. Андрогены оказывают влияние на появление оволосения на лобке и в подмышечных впадинах. Прогестерон контролирует секреторную фазу менструального цикла, подготавливает эндометрий к имплантации. Половые гормоны играют важную роль в процессе развития беременности и родов.
Циклические изменения в яичниках включают три основных процесса:
1. Рост фолликулов и формирование доминантного фолликула.
2. Овуляция.
3. Образование, развитие и регресс желтого тела.
При рождении девочки в яичнике находятся 2 млн фолликулов, 99 % которых подвергаются атрезии в течение всей жизни. Под процессом атрезии понимается обратное развитие фолликулов на одной из стадий его развития. Ко времени менархе в яичнике содержится около 200-400 тыс. фолликулов, из которых созревают до стадии овуляции 300-400.
Принято выделять следующие основные этапы развития фолликула (рис. 2.12): примордиальный фолликул, преантральный фолликул, антральный фолликул, преовуляторный фолликул.
Примордиальный фолликул состоит из незрелой яйцеклетки, которая расположена в фолликулярном и гранулезном (зернистом) эпителии. Снаружи фолликул окружен соединительной оболочкой (тека-клетки). В течение каждого менструального цикла от 3 до 30 примордиальных фолликулов начинают расти, и из них формируются преантральные, или первичные, фолликулы.
Преантральный фолликул. С началом роста примордиальный фолликул прогрессирует до преантральной стадии, а овоцит увеличивается и окружается мембраной, называемой блестящей оболочкой (zona pellucida). Клетки гранулезного эпителия подвергаются размножению, а слой теки образуется из окружающей стромы. Этот рост характеризуется повышением продукции эстрогенов. Клетки гранулезного слоя преантрального фолликула способны синтезировать стероиды трех классов, при этом эстрогенов синтезируется гораздо больше, чем андрогенов и прогестерона.
Антральный, или вторичный, ф о л л и к у л. Характеризуется дальнейшим ростом: увеличивается число клеток гранулезного слоя, продуцирующих фолликулярную жидкость. Фолликулярная жидкость накапливается в межклеточном пространстве гранулезного слоя и образует полости. В этот период фолликулогенеза (8-9-й день менструального цикла) отмечается синтез половых стероидных гормонов, эстрогенов и андрогенов.
Согласно современной теории синтеза половых гормонов, в тека-клетках синтезируются андрогены - андростендион и тестостерон. Затем андрогены попадают в клетки гранулезного слоя, и в них ароматизируются в эстрогены.
Доминантный фолликул. Как правило, один такой фолликул образуется из множества антральных фолликулов (к 8-му дню цикла). Он является самым крупным, содержит наибольшее число клеток гранулезного слоя и рецепторов к ФСГ, ЛГ. Доминантный фолликул имеет богато васкуляризированный тека-слой. Наряду с ростом и развитием доминантного преовуляторного фолликула в яичниках параллельно происходит процесс атрезии остальных (90 %) растущих фолликулов.
Доминантный фолликул в первые дни менструального цикла имеет диаметр 2 мм, который в течение 14 дней к моменту овуляции увеличивается в среднем до 21 мм. За это время происходит 100-кратное увеличение объема фолликулярной жидкости. В ней резко возрастает содержание эстрадиола и ФСГ, а также определяются факторы роста.
Овуляция - разрыв преовулярного доминантного (третичного) фолликула и выход из него яйцеклетки. Ко времени овуляции в овоците происходит процесс мейоза. Овуляция сопровождается кровотечением из разрушенных капилляров, окружающих тека-клетки. Полагают, что овуляция происходит через 24-36 ч после формирования преовуляторного пика эстрадиола. Истончение и разрыв стенки преовуляторного фолликула происходят под влиянием фермента коллагеназы. Определенную роль играют также простагландины F 2 a и Е 2 , содержащиеся в фолликулярной жидкости; протеолитические ферменты, образующиеся в гранулезных клетках; окситоцин и релаксин.
После выхода яйцеклетки в полость фолликула быстро врастают образующиеся капилляры. Гранулезные клетки подвергаются лютеинизации: в них увеличивается объем цитоплазмы и образуются липидные включения. ЛГ, взаимодействуя с белковыми рецепторами гранулезных клеток, стимулирует процесс их лютеинизации. Этот процесс приводит к образованию желтого тела.
Желтое тело - транзиторная эндокринная железа, которая функционирует в течение 14 дней независимо от продолжительности менструального цикла. При отсутствии беременности желтое тело регрессирует.
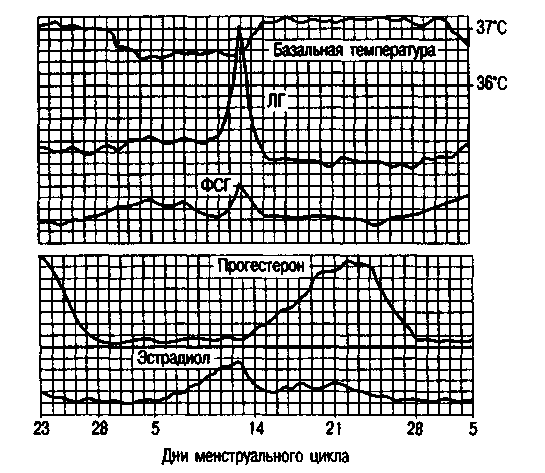
Рис. 2.13.
Содержание гормонов в плазме крови в течение менструального цикла.
Таким образом, в яичнике синтезируются основные женские половые стероидные гормоны - эстрадиол и прогестерон, а также андрогены.
В I фазу менструального цикла, который длится от первого дня менструации до момента овуляции, организм находится под влиянием эстрогенов, а во II (от овуляции до начала менструации) к эстрогенам присоединяется прогестерон, выделяющийся клетками желтого тела. Первая фаза менструального цикла называется также фолликулиновой, или фолликулярной, вторая фаза цикла - лютеиновой.
В течение менструального цикла в периферической крови отмечаются два пика содержания эстрадиола: первый -выраженный преовуляторный цикл, и второй, менее выраженный, -в середине второй фазы менструального цикла. После овуляции во второй фазе цикла основным является прогестерон, максимальное количество которого синтезируется на 4-7-й день после овуляции (рис. 2.13).
Циклическая секреция гормонов в яичнике определяет изменения в слизистой оболочке матки.
Циклические изменения в слизистой оболочке матки (эндометрии). Эндометрий состоит из следующих слоев.
1. Базальный слой, который не отторгается во время менструации. Из его клеток в течение менструального цикла образуется слой эндометрия.
2. Поверхностный слой, состоящий из компактных эпителиальных клеток, которые выстилают полость матки.
3. Промежуточный, или спонгиозный, слой.
Последние два слоя составляют функциональный слой, подвергающийся основным циклическим изменениям в течение менструального цикла и отторгающийся в период менструации.
В I фазе менструального цикла эндометрий представляет собой тонкий слой, состоящий из желез и стромы. Выделяют следующие основные фазы изменения эндометрия в течение цикла:
1) фаза пролиферации;
2) фаза секреции;
3) менструация.
Фаза пролиферации. По мере увеличения секреции эстрадиола растущими фолликулами яичников эндометрий претерпевает пролиферативные изменения. Происходит активное размножение клеток базального слоя. Образуется новый поверхностный рыхлый слой с вытянутыми трубчатыми железами. Этот слой быстро утолщается в 4-5 раз. Трубчатые железы, выстланные цилиндрическим эпителием, удлиняются.
Фаза секреции. В лютеиновую фазу яичникового цикла под влиянием прогестерона увеличивается извилистость желез, а просвет их постепенно расширяется. Клетки стромы, увеличиваясь в объеме, приближаются друг к другу. Секреция желез усиливается. В просвете желез находят обильное количество секрета. В зависимости от интенсивности секреции железы либо остаются сильно извитыми, либо приобретают пилообразную форму. Отмечается усиленная васкуляризация стромы. Различают раннюю, среднюю и позднюю фазы секреции.
Менструация. Это отторжение функционального слоя эндометрия. Тонкие механизмы, лежащие в основе возникновения и процесса менструации, неизвестны. Установлено, что эндокринной основой начала менструации является выраженное снижение уровней прогестерона и эстрадиола вследствие регрессии желтого тела.
Существуют следующие основные локальные механизмы, принимающие участие в менструации:
1) изменение тонуса спиральных артериол;
2) изменение механизмов гемостаза в матке;
3) изменения в лизосомной функции клеток эндометрия;
4) регенерация эндометрия.
Установлено, что началу менструации предшествует интенсивное сужение спиральных артериол, приводящее к ишемии и десквамации эндометрия.
В течение менструального цикла изменяется содержание лизосом в клетках эндометрия. Лизосомы содержат ферменты, некоторые из которых участвуют в синтезе простагландинов. В ответ на снижение уровня прогестерона усиливается выделение этих ферментов.
Регенерация эндометрия наблюдается с самого начала менструации. К концу 24-го часа менструации отторгается 2 /з функционального слоя эндометрия. Базальный слой содержит эпителиальные клетки стромы, являющиеся основой для регенерации эндометрия, которая обычно к 5-му дню цикла полностью завершается. Параллельно завершается ангиогенез с восстановлением целости разорванных артериол, вен и капилляров.
Изменения в яичниках и матке происходят под влиянием двухфазной деятельности регулирующих менструальную функцию систем: кора большого мозга, гипоталамус, гипофиз. Таким образом, выделяются 5 основных звеньев репродуктивной системы женщины: кора большого мозга, гипоталамус, гипофиз, яичник, матка (рис. 2.14). Взаимосвязь всех звеньев репродуктивной системы обеспечивается наличием в них рецепторов как к половым, так и к гонадотропным гормонам.
О роли ЦНС в регуляции функции репродуктивной системы известно давно. Об этом свидетельствовали нарушения овуляции при различных острых и хронических стрессах, нарушение менструального цикла при перемене климатогеографических зон, ритма работы; хорошо известно прекращение менструаций в условиях военного времени.У психически неуравновешенных женщин, страстно желающих иметь ребенка, менструации также могут прекратиться.
В коре большого мозга ив экстрагипоталамических церебральных структурах (лимбической системе, гиппокампе, миндалине и др.) выявлены специфические рецепторы для эстрогенов, прогестерона и андрогенов. В этих структурах происходят синтез, выделение и метаболизм нейропептидов, нейротрансмиттеров и их рецепторов, которые в свою очередь избирательно влияют на синтез и выделение рилизинг-гормона гипоталамуса.
Во взаимосвязи с половыми стероидами функционируют нейротрансмиттеры: норадреналин, дофамин, гамма-аминомасляная кислота, ацетилхолин, серотонин и мелатонин. Норадреналин стимулирует выброс гонадотропин-рилизинг-гормона (ГТРГ) из нейронов переднего гипоталамуса. Дофамин и серотонин уменьшают частоту и снижают амплитуду выработки ГТРГ в различные фазы менструального цикла.
Нейропептиды (эндогенные опиоидные пептиды, нейропептид Y, кортикотропин-рилизинг-фактор и галанин) также влияют на функцию репродуктивной системы, а следовательно, на функцию гипоталамуса. Эндоген- ные опиоидные пептиды трех видов (эндорфины, энкефалины и динорфины) способны связываться с опиатными рецепторами мозга. Эндогенные опиоидные пептиды (ЭОП) модулируют влияние половых гормонов на содержание ГТРГ по механизму обратной связи, блокируют секрецию гипофизом гонадотропных гормонов, особенно Л Г, посредством блокады секреции ГТРГ в гипоталамусе.
Взаимодействие нейротрансмитгеров и нейропептидов обеспечивает в организме женщины репродуктивного возраста регулярные овуляторные циклы, влияя на синтез и выделение ГТРГ гипоталамусом.
В гипоталамусе имеются пептидергические нейронные клетки, которые секретируют стимулирующие (либерины) и блокирующие (статины) нейрогормоны - нейросекреция. Эти клетки обладают свойствами как нейронов, так и эндокринных клеток, и отвечают как на сигналы (гормоны), поступающие из кровотока, так и на нейротрансмиттеры и нейропептиды мозга. Нейрогормоны синтезируются в рибосомах цитоплазмы нейрона, а затем транспортируются по аксонам к терминалям.
Гонадотропин-рилизинг-гормон (либерин) - нейрогормон, регулирующий гонадотропную функцию гипофиза, где синтезируются ФСГ и ЛГ. Рилизинг-гормон ЛГ (люлиберин) выделен, синтезирован и подробно описан. Выделить и синтезировать рилизинг-фолликулостимулирующий гормон, или фоллиберин, до настоящего времени не удалось.
Секреция гонадолиберина имеет пульсирующий характер: пики усиленной секреции гормона продолжительностью несколько минут сменяются 1- 3-часовыми интервалами относительно низкой секреторной активности. Частоту и амплитуду секреции гонадолиберина регулирует уровень эстрогенов.
Нейрогормон, который контролирует секрецию пролактина аденогипофизом, называется пролактинингибирующим гормоном (фактором), или дофамином.
Важным звеном в системе репродукции является передняя доля гипофиза - аденогипофиз, в котором секретируются гонадотропные гормоны, фолликулостимулирующий гормон (ФСГ, фоллитропин) лютеинизирующий гормон (ЛГ, лютропин) и пролактин (Прл), регулирующие функцию яичников и молочных желез. Все три гормона являются белковыми веществами (полипептидами). Железой-мишенью гонадотропных гормонов является яичник.
В передней доле гипофиза синтезируются также тиреотропный (ТТГ) и адренокортикотропный (АКТГ) гормоны, а также гормон роста.
ФСГ стимулирует рост и созревание фолликулов яичника, способствует образованию рецепторов ФСГ и ЛГ на поверхности гранулезных клеток яичника, увеличивает содержание ароматаз в зреющем фолликуле и, стимулируя процессы ароматизации, способствует превращению андрогенов в эстрогены, стимулирует продукцию ингибина, активина и инсулиноподобного фактора роста-1, которые играют ингибирующую и стимулирующую роль в росте фолликулов.
Л Г стимулирует:
Образование андрогенов в тека-клетках;
Овуляцию совместно с ФСГ;
Ремоделирование гранулезных клеток в процессе лютеинизации;
Синтез прогестерона в желтом теле.
Пролактин стимулирует рост молочных желез и лактацию, контролирует секрецию прогестерона желтым телом путем активации образования в них рецепторов к Л Г.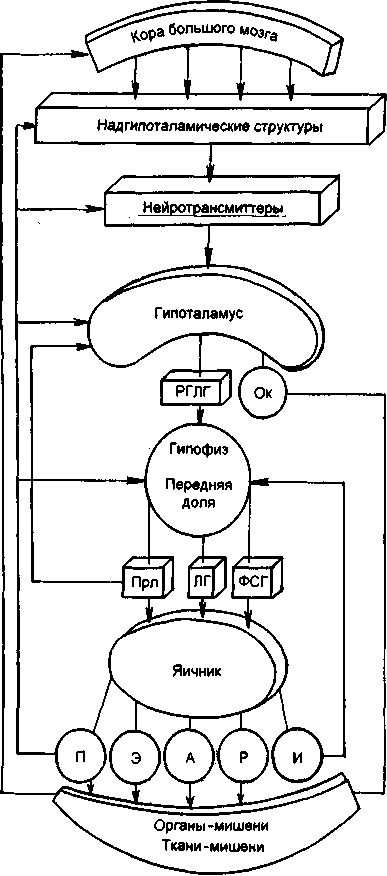
Рис. 2.14. Функция репродуктивной системы (схема).
РГЛГ - рилизинг-гормоны; ОК - окситоцин; Прл - пролактин; ФСГ - фолликуло-стимулирующий гормон; П - прогестерон; Э - эстрогены; А - андрогены; Р - релаксин; И - ингибин; ЛГ - лютеинизирующий гормон.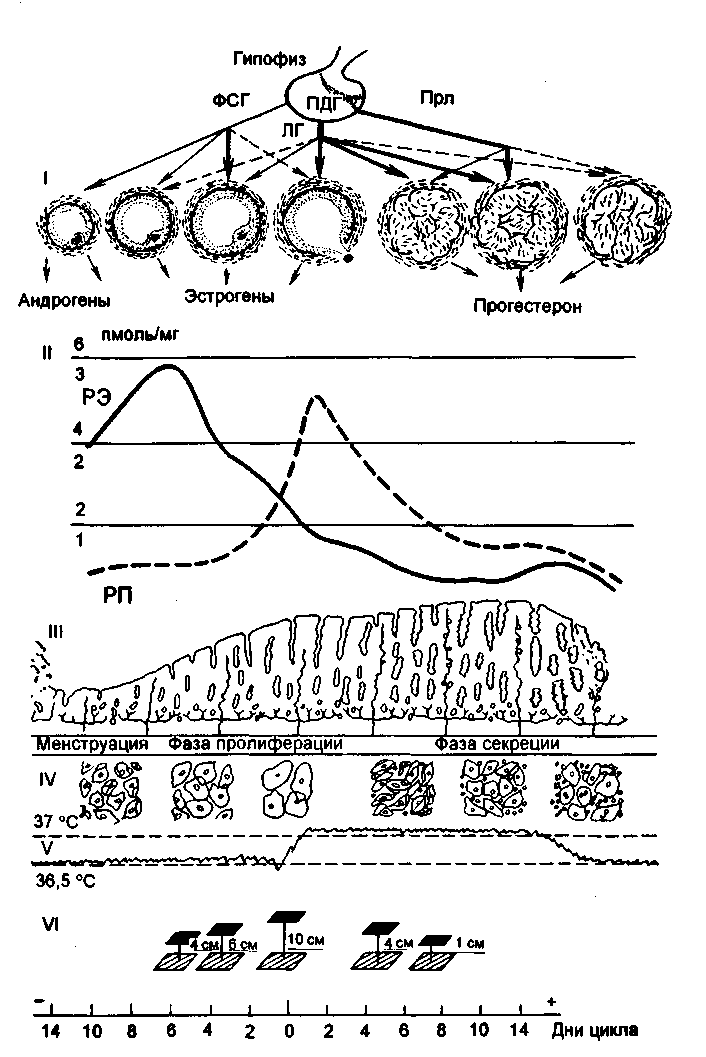
Рис. 2.15.
Циклические изменения в органах репродуктивной системы в течение менструального цикла. I - гонадотропная регуляция функции яичников: ПДГ - передняя доля гипофиза, остальные обозначения те же, что на рис. 2.14; II - содержание в эндометрии рецепторов к эстрадиолу - РЭ (1,2,3; сплошная линия) и прогестерону - РП (2,4,6; пунктирная линия); III - циклические изменения эндометрия; IV - цитология эпителия влагалища; V - базальная температура; VI - натяжение цервикальной слизи.
Синтез пролактина аденогипофизом находится под тоническим блокирующим контролем дофамина, или пролактинингибирующего фактора. Ингибиция синтеза пролактина прекращается во время беременности, лактации. Основным стимулятором синтеза пролактина является тиролиберин, синтезируемый в гипоталамусе.
Циклические изменения в гипоталамо-гипофизарной системе и в яичниках взаимосвязаны и моделируются по типу обратной связи.
Выделяют следующие типы обратной связи:
1) "длинная петля" обратной связи - между гормонами яичника и ядрами гипоталамуса; между гормонами яичника и гипофизом;
2) "короткая петля" - между передней долей гипофиза и гипоталамусом;
3) "ультракороткая петля" - между ГТРГ и нервными клетками гипоталамуса.
Взаимосвязь всех указанных структур определяется наличием в них рецепторов к половым гормонам.
У женщины репродуктивного возраста имеется как отрицательная, так и положительная обратная связь между яичниками и гипоталамо-гипофизарной системой. Примером отрицательной обратной связи является усиление выделения ЛГ передней долей гипофиза в ответ на низкий уровень эстрадиола в раннюю фолликулярную фазу цикла. Примером положительной обратной связи является выброс ЛГ в ответ на овуляторный максимум содержания эстрадиола в крови.
О состоянии репродуктивной системы можно судить по оценке тестов функциональной диагностики: базальная температура, симптом зрачка и кариопикнотический индекс (рис. 2.15).
Базальная температура измеряется в прямой кишке утром, до подъема с постели. При овуляторном менструальном цикле базальная температура повышается в лютеиновую фазу цикла на 0,4-0,6 °С и держится в течение всей второй фазы (рис. 2.16). В день менструации или за день до нее базальная температура снижается. При беременности повышение базальной температуры объясняется возбуждением терморегулирующего центра гипоталамуса под влиянием прогестерона.
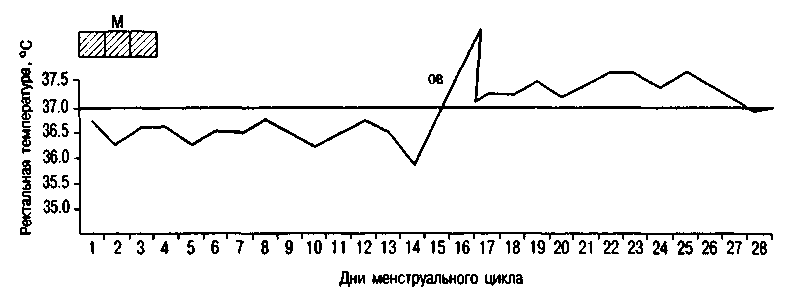
Рис. 2.16. Ректальная температура при двухфазном цикле. М - менструация; ОВ - овуляция.
Симптом зрачка отражает изменения в слизи шейки матки. Под влиянием эстрогенов в шейке матки накапливается прозрачная стекловидная слизь, что обусловливает расширение наружного отверстия шейки матки. Максимальное количество слизи наблюдается в предовуляторные дни цикла, наружное отверстие становится темным, напоминает зрачок. Во вторую фазу цикла под влиянием прогестерона количество слизи уменьшается или же она полностью исчезает. Слизь имеет глыбчатое строение. Различают 3 степени симптома зрачка: +, ++, +++.
Кариопикнотический индекс. Под влиянием гормонов яичников также происходят циклические изменения в слизистой оболочке влагалища, особенно в его верхней трети. В мазке из влагалища могут встречаться следующие виды клеток плоского многослойного эпителия: а) ороговевающие, б) промежуточные, в) базальные, или атрофические. Клетки первого типа начинают преобладать по мере нарастания секреции яичниками эстрогенов. На основании определения количественных соотношений клеточных элементов можно судить о степени насыщенности организма эстрогенными гормонами или об их недостаточности. Максимальное число ороговевающих клеток выявляется в предовуляторные дни - 80-88 %, в раннюю фазу пролиферации - 20-40 %, в позднюю фазу секреции - 20-25 %.
Этот пример иллюстрирует основной способ преобразования энергии в
клетке : химическая работа совершается путем подключения к реакции с
«неблагоприятным» изменением свободной энергии реакций с большим
отрицательным изменением свободной энергии. Чтобы осуществлять
такое «сопряжение» процессов, клетке пришлось создать в ходе эволюции
специальные молекулярные «энергопреобразующие» устройства, которые
представляют собой ферментные комплексы, как правило, связанные с
мембранами.
Механизмы трансформации энергии в биоструктурах связаны с конформационными превращениями особых макромолекулярных комплексов, таких, как реакционные центры фотосинтеза, Н – АТФаза хлоропластов и митохондрий, бактериородопсин. Особый интерес представляют общие характеристики эффективности преобразования энергии в таких макромолекулярных машинах. На эти вопросы призвана ответить термодинамика биологических процессов.
Женские половые органы делятся на наружные и внутренние.
Наружные половые органы.
К наружным половым органам у женщин относятся: лобок,большие и малые половые губы,бартолиновы железы,клитор,преддверие влагалища и девственная плева,которая является границей между наружными и внутренними половыми органами.
ЛОБОК – треугольное возвышение,покрытое волосами,распологающееся над лоном. Границами являются: сверху – поперечная кожная борозда;с боков – паховые складки.
У женщин верхняя граница волосистого покрова лобка имеет вид горизонтальной линии.
БОЛЬШИЕ ПОЛОВЫЕ ГУБЫ – две кожные складки ограничивающие с боков половую щель.Спереди переходят в кожу лобка,кзади сливаются в заднюю спайку.Кожа на наружной поверхности больших половых губ покрыта волосами,содержит потовые и сальные железы,под ней в подкожно-жировой клетчатке залегают сосуды,нервы и фиброзные волокна,а в задней трети – большие железы преддверия (бартолиновы железы) – округлые альвеолярно-трубчатые,
величиной с боб железы.Их выводные протоки открываются в борозде между малыми половыми губами и девственной плевой,а их секрет выделяется при половом возбуждении.
Пространство между задней спайкой и заднепроходным отверстием называется промеж-
ностью.В анатомическом смысле промежность представляет собой мышечно-фасциальную пластинку,покрытую снаружи кожей.Высота её в среднем равна 3-4 см.
МАЛЫЕ ПОЛОВЫЕ ГУБЫ – вторая пара продольных кожных складок.Располагаются кнутри от больших половых губ и обычно прикрыты последними.Спереди малые половые губы раздваиваются на две ножки с каждой стороны,которые сливаясь,образуют крайнюю плоть клитора и уздечку клитора.Кзади малые половые губы сливаются с большими.Благодаря оби-
лию сосудов и нервных окончаний малые половые губы являются органами полового чувства.
КЛИТОР. Внешне заметен как небольшой бугорок в переднем углу половой щели между слившимися ножками малых половых губ.В клиторе различают головку,тело,состоящее из пещеристых тел и ножки,которые прикрепляются к надкостнице лобковых и седалищных костей.Обильное кровоснабжение и иннервация делают его основным органом полового чувства женщины.
ПРЕДДВЕРИЕ ВЛАГАЛИЩА – пространство,ограниченное спереди клитором,сзади задней спайкой половых губ,с боков – внутренней поверхностью малых половых губ,сверху – девственной плевой.Сюда открываются наружное отверстие уретры и выводные протоки бартолиновых желез.
ДЕВСТВЕННАЯ ПЛЕВА – соединительно-тканная перепонка,закрывающая вход во влагалище у девственниц.В её соединительнотканной основе имеются мышечные элементы, кровеносные сосуды и нервы.В дественной плеве должно быть отверстие.Оно может быть любой формы.После дефлорации от плевы остаются гименальные сосочки,после родов – миртовидные сосочки.
Внутренниеполовые органы.
К ним относятся:влагалище,матка,маточные трубы и яичники.
ВЛАГАЛИЩЕ – хорошо растяжимая,мышечно-эластичная трубка.Идет спереди и снизу кзади и кверху.Начинается от девственной плевы и заканчивается у места прикрепления к шейке матки.Средние размеры:длина 7-8 см. (задняя стенка на 1,5-2 см. длиннее),ширина 2-3 см.Благодаря тому,что передняя и задняя стенки влагалища соприкасаются,на поперечном разрезе оно имеет форму буквы Н. Вокруг влагалищной части шейки матки,которая выступает во влагалище,стенки влагалища образуют сводчатое образование.Его принято делить на передний,задний (наиболее глубокий) и боковые своды.Стенка влагалища состоит из трёх слоёв:слизистого,мышечного и окружающей клетчатки,в которой проходят сосуды и нервы.Мышечный слой состоит из двух слоёв:наружного продольного и внутреннего циркулярного.Слизистая покрыта многослойным плоским эпителием,содержащим гликоген. Процесс образования гликогена связан с фолликулярным гормоном яичника.Влагалище очень хорошо растяжимо благодаря наличию на передней и задней стенках двух продольных валиков,состоящих из множества поперечных складок.В слизистой влагалища нет желез.Секрет влагалища образуется за счёт пропитывания жидкости из сосудов.Он имеет кислую среду благодаря образующейся из гликогена под влиянием ферментов и продуктов жизнедеятельности лактобацилл (палочек Дедерлейна) молочной кислоты.Молочная кислота способствует гибели патогеннных микроорганизмов.
Различают четыре степени чистоты влагалищного содержимого.
1 степень : в содержимом только лактобациллы и эпителиальные клетки,реакция кислая.
2 степень : палочек Дедерлейна меньше,единичные лейкоциты,бактерии,много эпителиальных клеток,реакция кислая.
3 степень : лактобацилл мало,преобладают другие виды бактерий,много лейкоцитов,реакция слабощелочная.
4 степень : лактобацилл нет,очень много бактерий и лейкоцитов,реакция щелочная.
1,2 степени – вариант нормы.
3,4 степени говорят о наличии патологического процесса.
МАТКА – гладкомышечный полый орган,имеющий форму груши,уплощённый в переднезаднем направлении.
Отделы матки:тело,перешеек,шейка.
Куполообразно возвышающаяся часть тела выше линий прикрепления труб называется дном матки.
Перешеек – часть матки длиной 1 см,располагающаяся между телом и шейкой.Выделяется в отдельный отдел,так как по строению слизистой похож на тело матки,а по строению стенки на шейку матки.Верхней границей перешейка является место плотного прикрепления брюшины к передней стенке матки.Нижней границей является уровень внутреннего зева канала шейки матки.
Шейка – нижняя часть матки,выступающая во влагалище.В ней различают две части:влага-лищную и надвлагалищную.Шейка матки может быть либо цилиндрической,либо конической формы (детский возраст,инфантилизм).Внутри шейки матки расположен узкий канал,имеющий веретёнообразную форму,ограниченный внутренним и наружным зевами.Наружный зев открывается в центре влагалищной части шейки.Он имеет форму щели у рожавших и округлую форму у нерожавших женщин.
Длина всей матки 8 см. (2/3 длины приходится на тело,1/3 – на шейку),ширина 4-4,5 см, толщина стенок 1-2 см.Масса 50-100г.Полость матки имеет форму треугольника.
Стенка матки состоит из 3 слоёв:слизистого,мышечного,серозного.Слизистая оболочка матки (эндометрий) покрыта однослойным цилиндрическим мерцательным эпителием,содержащим трубчатые железы.Слизистая матки делится на два слоя:поверхностный (функционвльный),от-торгающийся во время менструации,глубокий (базальный),остающийся на месте.
Мышечный слой (миометрий) богато снабжён сосудами,состоит из трёх мощных пластов:на-ружного продольного;среднего циркулярного;внутреннего продольного.
Серозный покров матки (периметрий) – это брюшина,покрывающая тело и частично шейку матки.С мочевого пузыря брюшина переходит на переднюю поверхность матки,образуя между двумя этими органами пузырно-маточное углубление.С дна матки брюшина спускается по задней её поверхности,выстилая надвлагалищную часть шейки матки и задний свод влагалища,а затем переходит на переднюю поверхность прямой кишки,таким образом образуя глубокий карман – прямокишечно-маточное углубление (дугласово пространство).
Матка находится в центре малого таза,наклонена кпереди (anteversio uteri),дно её направлено к симфизу,шейка кзади,наружный зев шейки примыкает к стенке заднего свода влагалища.Между телом и шейкой матки существует тупой угол,открытый кпереди (anteflexio uteri).
МАТОЧНЫЕ ТРУБЫ начинаются от верхних углов матки,идут по верхнему краю широкой связки в сторону боковых стенок таза,заканчиваясь воронкой.Длина их 10-12 см.В трубе различают три отдела:1) интерстициальный – самая узкая часть,проходящая в толще матки;2) перешеечный (истмус); 3) ампулярный – расширенная часть трубы,заканчивающаяся воронкой с фимбриями.В этом отделе трубы происходит оплодотворение – слияние яйцеклетки и сперматозоида.
Стенка труб состоит из трёх слоёв:слизистого,мышечного,серозного.
Слизистая покрыта однослойным цилиндрическим мерцательным эпителием,имеет продольную складчатость.
Мышечный слой состоит из трёх пластов: наружного – продольного;среднего – циркулярного;внутреннего – продольного.
Брюшина покрывает трубу сверху и с боков.К нижнему отделу трубы прилегает клетчатка с сосудами и нервами.
Продвижению по трубе оплодотворённой яйцеклетки в сторону матки способствуют перестальтические сокращения мускулатуры трубы, мерцание ресничек эпителия, направленное в сторону матки и продольная складчатость слизистой трубы. По складке как по жёлобу яйцеклетка скользит по направлению к матке.
ЯИЧНИКИ – парная женская гонада миндалевидной формы, размерами 3,5-4 х 2-2,5 х 1-1,5 см, массой 6-8г.
Яичник одним краем вставлен в задний листок широкой связки (ворота яичника),вся остальная его часть брюшиной не покрыта.Яичник удерживается в свободно подвешенном состоянии широкой маточной связкой, собственной связкой яичника, воронкотазовой связкой.
В яичнике различают покровный эпителий, белочную оболочку, корковый слой с фолликулами на разных стадиях развития, мозговой слой, состоящий из соединительно-тканной стромы, в которой проходят сосуды и нервы.
В яичниках вырабатываются половые гормоны, созревают яйцеклетки.
Связочный аппарат половых органов.
В нормальном положении матку с придатками удерживают связочный аппарат (подвешивающий и закрепляющий аппараты) и мышцы тазового дна (поддерживающий или опорный аппарат).
В подвешивающий аппарат входят:
1.Круглые маточные связки – два шнура длиной 10-12 см.Отходят от маточных углов,и пройдя под широкой маточной связкой и через паховые каналы,веерообразно разветвляются, прикрепляясь к клетчатке лобка и больших половых губ.
2.Широкие связки матки – дубликатура брюшины.Идут от рёбер матки к боковым стенкам таза.
3.Крестцово-маточные связки – отходят от задней поверхности матки в области перешейка,идут
кзади,охватывая с двух сторон прямую кишку.Прикрепляются на передней поверхности крестца.
4.Собственные связки яичников идут от дна матки (кзади и ниже места отхождения труб) к яичникам.
5.Воронкотазовые связки – самая наружная часть широкой маточной связки,переходящая в брюшину боковой стенки таза.
Круглые связки удерживают матку в состоянии anteversiо,широкие связки при перемещении матки напрягаются и тем самым способствуют удержанию матки в физиологическом положении,собственные связки яичника и воронкотазовые связки способствуют удержанию матки в срединном положении,крестцово-маточные связки притягивают матку кзади.
Закрепляющий аппарат матки состоит из соединительнотканных тяжей с небольшим количеством мышечных клеток,которые идут от нижнего отдела матки: а)кпереди к мочевому пузырю и далее к симфизу; б)к боковым стенкам таза – основные связки; в)кзади,составляя соединительнотканный остов крестцово-маточных связок.
Поддерживающий аппарат составляют мышцы и фасции тазового дна,которые препятствуют опусканию вниз половых органов и внутренностей.
Кровоснабжение половых органов.
Наружные половые органы кровоснабжаются за счёт срамной артерии (веточка внутренней подвздошной артерии).
Кровоснабжение внутренних половых органов обеспечивается маточной и яичниковой артериями.
Маточная артерия парная,отходит от внутренней подвздошной артерии,идёт к матке по околоматочной клетчатке,подойдя к боковой поверхности матки на уровне внутреннего зева, отдаёт шеечно-влагалищную ветвь,кровоснабжающую шейку и верхний отдел влагалища. Основной же ствол поднимается по ребру матки,отдавая многочисленные веточки,питающие стенку матки, и доходит до дна матки,где отдаёт веточку,идущую к трубе.
Яичниковая артерия тоже парная,отходит от брюшной аорты,спускается вниз вместе с моче-точником,проходит по воронкотазовой связке,отдавая ветки к яичнику и трубе.Конечные отделы маточной и яичниковой артерий анастомозируют между собой в верхнем отделе широкой маточной связки.
Артерии сопровождаются одноимёнными венами.
Иннервация половых органов.
В иннервации половых органов принимают участие симпатическая и парасимпатическая нервные системы (маточно-влагалищное и яичниковое сплетения).
Наружные половые органы и тазовое дно иннервируются срамным нервом.
Физиология женских половых органов.
Известно,что репродукция,или воспроизводство,является одной из важнейших функций
женского организма.Репродуктивная функция женщин осуществляется прежде всего благодаря деятельности яичников и матки,так как в яичниках созревает яйцеклетка,а в матке под влиянием гормонов,выделяемых яичниками,происходят изменения по подготовке к восприятию оплодо- творённого плодного яйца.Репродуктивный (детородный) период продолжается с 17-18 до 45-50 лет.
Детородному периоду предшествуют следующие этапы жизни женщины:внутриутробный; новорожденности (до1 года);детства (до 8-10 лет);препубертатного и пубертатного возраста (до 17-18 лет).Репродуктивный период переходит в климактерический,в котором различают пременопаузу,менопаузу (последняя менструация) и постменопаузу.
Менструальный цикл – одно из проявлений сложных биологических процессов в организме женщины.Менструальный цикл характеризуется циклическими изменениями во всех звеньях репродуктивной системы,внешним проявлением которых является менструация.
Каждый нормальный менструальный цикл является подготовкой организма женщины к беременности.Зачатие и беременность наступают обычно в середине менструального цикла после овуляции (разрыв зрелого фолликула) и выхода из яичника готовой к оплодотворению яйцеклетки.Если в этот период оплодотворение не происходит,неоплодотворённая яйцеклетка погибает,а подготовленная для её восприятия слизистая оболочка матки отторгается и начинается менструальное кровотечение.Таким образом,появление менструации свидетельствует об окончании сложных циклических изменений в организме женщины, направленных на подготовку к возможному наступлению беременности.
Первый день менструации условно принимается за первый день менструального цикла,а про-должительность цикла определяют от начала одной и до начала другой (последующей) менструации.Нормальная продолжительность менструального цикла колеблется от 21 до 35 дней и у большинства женщин в среднем составляет 28 дней.Величина кровопотери в менструальные дни 50-100 мл.Продолжительность нормальной менструации от 2 до 7 дней.
Первая менструация (menarhe) наблюдается в возрасте 10-12 лет,но в течение 1-1,5 года после этого менструации могут быть нерегулярными,затем устанавливается регулярный менструальный цикл.
Регуляция менструальной функции осуществляется сложным нейрогуморальным путём при участии пяти звеньев (уровней):1)кора головного мозга;2)гипоталамус;3)гипофиз;4)яичники; 5)периферические органы,называемые органами-мишенями (маточные трубы,матка и влагалище).Органы-мишени,благодаря наличию особых гормональных рецепторов наиболее чётко реагируют на действие половых гормонов,вырабатываемых в яичниках во время менструального цикла.
Циклические функциональные изменения,происходящие в организме женщины,условно объединены в несколько групп.Это изменения в системе гипоталамус-гипофиз,яичниках (яичниковый цикл),матке и в первую очередь в её слизистой оболочке (маточный цикл).Наряду с этим происходят циклические сдвиги во всём организме женщины,известные под названием менструальной волны.Они выражаются в периодических изменениях деятельности ЦНС, обменных процессов,функции ССС-мы,терморегуляции и др.
Кора головного мозга осуществляет регулирующее и корригирующее влияние на процессы,связанные с развитием менструальной функции.Через кору головного мозга осуществляется влияние внешней среды на нижележащие отделы нервной системы, участвующие в регуляции менструального цикла.
Гипоталамус является отделом промежуточного мозга и при помощи ряда нервных проводников (аксонов) соединён с различными отделами головного мозга,благодаря чему осуществляется центральная регуляция его активности.Кроме того,гипоталамус содержит рецепторы для всех периферических гормонов,в том числе яичниковых (эстрогенов и прогестерона).Таким образом,в гипоталамусе осуществляются сложные взаимодействия между импульсами,поступающими в организм из окружающей среды через ЦНС,с одной стороны, и
влияниями гормонов переферических желез внутрнней секреции – с другой.
Под контролем гипоталамуса находится деятельность придатка мозга –гипофиза,в передней доле которого выделяются гонадотропные гормоны,оказывающие влияние на функцию яичников.
Контролирующее действие гипоталамуса на переднюю долю гипофиза осуществляется посредством секреции нейрогормонов.
Нейрогормоны,стимулирующие освобождение тропных гормонов гипофиза,называются рилизинг-факторами или либеринами.Наряду с этим существуют также нейрогормоны, ингибирующие освобождение тропных нейрогормонов, называемые статинами.
В передней доле гипофиза секретируются фолликулостимулирующий (ФСГ) и лютеинизирующий (ЛТ) гонадотропины,а также пролактин.
ФСГ стимулирует развитие и созревание фолликула в одном из яичников.Под комбинированным воздействием ФСГ и ЛГ происходит разрыв зрелого фолликула,или овуляция.После овуляции под преобладающим воздействием ЛГ из элементов фолликула (соединительнотканной оболочки и выстилающих его внутреннюю поверхность гранулёзных клеток) образуется жёлтое тело.Пролактин способствует продукции жёлтым телом гормона прогестерона.
В яичниках во время менструального цикла происходит рост фолликулов и созревание яйцеклетки,которая в результате становится готовой к оплодотворению.Одновременно в яичниках вырабатываются половые гормоны,обеспечивающие изменения в слизистой оболочке матки,способной воспринять оплодотворённое яйцо.
Половые гормоны,синтезируемые яичниками,влияют на ткани- и органы-мишени, взаимодействуя с соответствующими рецепторами.К тканям- и органам-мишеням относятся половые органы,в первую очередь матка,молочные железы,губчатое вешество костей, мозг,эндотелий и гладкие мышечные клетки сосудов,миокард,кожа и её придатки (волосяные фолликулы и сальные железы) и др.
Эстрогенные гормоны способствуют формированию половых органов,развитию вторичных половых признаков в период полового созревания.Андрогены оказывают влияние на появление оволосения на лобке и в подмышечных впадинах.Прогестерон контролирут секреторную фазу менструального цикла,подготавливает эндометрий к имплантации.Половые гормоны играют важную роль в развитии беременности и родов.
Циклические изменения в яичниках включают три основных процесса:
1)рост фолликулов и формирование доминантного фолликула (фолликулярная фаза);
2)овуляцию;
3)образование,развитие и регресс жёлтого тела (лютеиновая фаза).
При рождении девочки в яичнике находится 2 млн фолликулов,99% которых подвергаются атрезии в течение всей жизни.Под процессом атрезии понимается обратное развитие фолликулов на одной из стадий его развития.Ко времени менархе в яичнике содержится около 200-400 тыс. фолликулов,из которых созревают до стадии овуляции 300-400.
Принято выделять следующие основные этапы развития фолликула:примордиальный фолликул,преантральный фолликул,антральный фолликул,преовуляторный (доминантный) фолликул.Доминантный фолликул является самым крупным (к моменту овуляции 21 мм).
Овуляция –разрыв доминантного фолликула и выход из него яйцеклетки.Истончение и разрыв стенки фолликула происходит в основном под влиянием фермента коллагеназы.
После выхода яйцеклетки в полость фолликула быстро врастают образующиеся капилляры.Гранулёзные клетки подвергаются лютеинизации:в них увеличивается объём цитоплазмы и образуются липидные включения.ЛГ,взаимодействуя с белковыми рецепторами гранулёзных клеток,стимулирует процесс их лютеинизации.Этот процесс приводит к образованию жёлтого тела.
Жёлтое тело –транзиторная эндокринная железа,которая функционирует в течение 14 дней независимо от продолжительности менструального цикла.При отсутствии беременности жёлтое тело регрессирует.
Циклическая секреция гормонов в яичнике определяет изменения в слизистой оболочке матки. Эндометрий состоит из двух слоёв: базального,который не отторгается во время менструации и функционального,подвергающегося циклическим изменениям в течение менструального цикла и отторгающегося в период менструации.
Выделяют следующие фазы изменения эндометрия в течение цикла:
1)фаза пролиферации; 3)менструация;
2)фаза секреции; 4)фаза регенерации
Фаза пролиферации. По мере увеличения секреции эстрадиола растущими фолликулами яичников эндометрий претерпевает пролиферативные изменения.Происходит активное размножение клеток базального слоя.Образуется новый поверхностный рыхлый слой с вытянутыми трубчатыми железеми.Этот слой быстро утолщается в 4-5 раз.Трубчатые железы,выстланные цилиндрическим эпителием,удлиняются.
Фаза секреции. В лютеиновую фазу яичникового цикла под влиянием прогестерона увеличивается извилистость желез,а их просвет постепенно расширяется.Клетки стромы, увеличиваясь в объёме,приближаются друг к другу.Секреция желез усиливается.Они приобретают пилообразную форму.Отмечается усиленная васкуляризация стромы.
Менструация. Это отторжение функционального слоя эндометрия.Эндокринной основой начала менструации является выраженное снижение уровней прогестерона и эстрадиола вследствие регрессии жёлтого тела.
Фаза регенерации. Регенерация эндометрия наблюдается с самого начала менструации.К концу 24-го часа менструации отторгается 2/3 функционального слоя эндометрия.Базальный слой содержит эпителиальные клетки стромы,являющиеся основой для регенерации эндометрия,которая обычно к 5-му дню цикла полностью завершается.Параллельно завершается ангиогенез с восстановлением целости разорванных артериол,вен и капиляров.
В регуляции менструальной функции большое значение имеет осуществление принципа так называемой обратной связи между гипоталамусом,передней долей гипофиза и яичниками.Принято рассматривать два типа обратной связи:негативную и позитивную.
При негативном типе обратной связи продукция центральных нейрогормонов (рилизинг-факторов) и гонадотропинов аденогипофиза подавляется гормонами яичников, вырабатыва- емыми в больших количествах.При позитивном типе обратной связи выработка рилизинг-факторов в гипоталамусе и гонадотропинов в гипофизе стимулируется низким содержанием в крови яичниковых гормонов.Осуществление принципа негетивной и позитивной обратной связи лежит в основе саморегуляции функции системы гипоталамус -гипофиз-яичники.
Женский таз и тазовое дно.
Костный таз имеет большое значение в акушерстве.Он представляет собой вместилище для внутренних половых органов, прямой кишки, мочевого пузыря и окружающих их тканей, а во время родов образует родовой канал по которому происходит продвижение плода.
Таз состоит из четырёх костей: двух тазовых (безымянных),крестца и копчика.
Тазовая кость состоих из трёх костей: подвздошной,лобковой и седалищной,соединённых между собой в области вертлужной впадины.
Различают два отдела таза: большой таз и малый таз. Граница между ними проходит спереди по верхнему краю лонного сочленения, с боков по безымянной линии, сзади по крестцовому мысу.
Большой таз ограничен с боков крыльями подвздошных костей, сзади – последними поясничными позвонками. Спереди он не имеет костной стенки. По размерам большого таза, которые достаточно легко измерить судят о форме и размерах малого таза.
Малый таз представляет собой костную часть родового канала. Форма и размеры малого таза имеют большое значение в течении родового акта. При резких степенях сужения таза и его деформациях роды через естественные родовые пути становятся невозможными, и женщину родоразрешают путём операции кесарева сечения.
Задняя стенка малого таза состоит из крестца и копчика, боковые образованы седалищными костями,передняя - лобковыми костями и симфизом. Задняя стенка малого таза в три раза длиннее передней.
В малом тазу существуют следующие отделы: вход, полость и выход. В полости таза различают широкую и узкую часть. В соответствии с этим рассматривают четыре плоскости малого таза:1)плоскость входа в малый таз;2)плоскость широкой части малого таза;3)плоскость узкой части малого таза;4)плоскость выхода таза.
Плоскость входа в таз имеет следующие границы: спереди – верхний край симфиза и лобковых костей, с боков – безымянные линии, сзади – крестцовый мыс. Плоскость входа имеет почкообразную форму. В плоскости входа различают следующие размеры: прямой, являющийся истинной конъюгатой малого таза (11 см),поперечный (13 см) и два косых (12 см).
Плоскость широкой части полости таза ограничена впереди серединой внутренней поверхности симфиза, по бокам- серединой вертлужных впадин, сзади- местом соединения II и III крестцовых позвонков.В широкой части различают два размера прямой (12,5 см) и поперечный (12,5 см)
Плоскость узкой части полости таза ограничена спереди нижним краем симфиза, с боков остями седалищных костей, сзади крестцово-копчиковым соединением. Здесь также два размера: прямой (11 см) и поперечный (10,5 см).
Плоскость выхода таза имеет следующие границы: спереди – нижний край симфиза, с боков – седалищные бугры, сзади –копчик. Плоскость выхода таза состоит из двух треугольных плоскостей, общим основанием которых является линия, соединяющая седалищные бугры. Прямой размер выхода таза – от верхушки копчика до нижнего края симфиза, за счёт подвижности копчика при прохождении плода через малый таз увеличивается на 1,5 – 2 см (9,5-11,5 см). Поперечный размер равен 11 см.
Линия, соединяющая середины прямых размеров всех плоскостей таза называется проводной осью таза , так как именно по этой линии во время родов плод проходит через родовой канал. Проводная ось изогнута соответственно вогнутости крестца.
Пересечение плоскости входа в таз с плоскостью горизонта образует угол наклонения таза равный 50-55’.
Отличия в строении женского и мужского таза начинают проявляться в период полового созревания и становятся выраженными в зрелом возрасте. Кости женского таза более тонкие, гладкие и менее массивные, чем кости мужского таза. Плоскость входа в малый таз у женщин имеет поперечно-овальную форму, в то время как у мужчин она имеет форму карточного сердца (вследствие сильного выступания мыса).
В анатомическом отношении женский таз ниже, шире и больше в обёме. Лобковый симфиз в женском тазу короче мужского. Крестец у женщин шире, крестцовая впадина умеренно вогнута. Полость малого таза у женщин по очертаниям приближается к цилиндру, а у мужчин воронкообразно сужается книзу. Лобковый угол шире (90-100’),чем у мужчин (70-75’).Копчик выдаётся кпереди меньше, чем в мужском тазу. Седалищные кости в женском тазу параллельны друг другу, а в мужском сходятся.
Все перечисленные особенности имеют очень большое значение в процессе родового акта.
Мышцы тазового дна.
Выход таза закрыт снизу мощным мышечно-фасциальным пластом, который называется тазовым дном.
В образовании дна малого таза принимают участие две диафрагмы – тазовая и мочеполовая.
Диафрагма таза занимает заднюю часть промежности и имеет вид треугольника, вершина которого обращена к копчику, а углы – к седалищным буграм.
Поверхностный слой мышц диафрагмы таза представлен непарной мышцей – наружным сфинктером заднего прохода (m.sphincter ani externus).Глубокие пучки этой мышцы начинаются от верхушки копчика, обхватывают заднепроходное отверстие и оканчиваются в сухожильном центре промежности.
К глубоким мыщцам диафрагмы таза относятся две мышцы: мышца, поднимающая задний проход (m.levator ani) и копчиковая мышца (m. coccygeus).
Мышца, поднимающая задний проход –парная, треугольной формы, образует с аналогичной мышцей другой стороны воронку, широкой частью, обращённую кверху и прикрепляющуюся к внутренней поверхности стенок таза. Нижние части обеих мышц, суживаясь, охватывают прямую кишку в виде петли. Эта мышца состоит из лобково-копчиковой (m. pubococcygeus) и подвздошно-копчиковой мышц (m.iliococcygeus).
Копчиковая мышца в виде треугольной пластинки располагается на внутренней поверхности крестцово-остистой связки. Узкой верхушой она начинается от седалищной ости, широким основанием прикрепляется к боковым краям нижних крестцовых и копчиковых позвонков.
Мочеполовая диафрагма –фасциально –мышечная пластинка, расположенная в передней части дна малого таза между нижними ветвями лобковых и седалищных костей.
Мышцы мочеполовой диафрагмы делятся на поверхностные и глубокие.
К поверхностным относятся поверхностная поперечная мышца промежности, седалищно-пещеристая мышца и луковично-губчатая.
Поверхностная поперечная мышца промежности (m.transversus perinei superficialis) – парная,непостоянная, иногда может отсутствовать на одной или обеих сторонах. Эта мышца представляет тонкую мышечную пластинку, расположенную у заднего края мочеполовой диафрагмы и идущую поперёк промежности. Латеральным концом она прикрепляется к седалищной кости,медиальной частью перекрещивается по средней линии с одноимённой мышцей противоположной стороны, частично вплетаясь в луковично-губчатую мышцу, частично – в наружную мышцу, сжимающую задний проход.
Седалищно-пещеристая мышца (m.ischiocavernosus) – парная, имеющая вид узкой мышечной полоски. Она начинается узким сухожилием от внутренней поверхности седалищного бугра, обходит ножку клитора и вплетается в его белочную оболочку.
Луковично-губчатая мышца (m. bulbospongiosus) – парная, окружает вход во влагалище, имеет форму вытянутого овала. Эта мышца начинается от сухожильного центра промежности и наружного сфинктера заднего прохода и прикрепляется к дорсальной поверхности клитора, вплетаясь в его белочную оболочку.
К глубоким мышцам мочеполовой диафрагмы относятся глубокая поперечная мышца промежности и сфинктер мочеиспускательного канала.
Глубокая поперечная мышца промежности (m. transversus perinei profundus) – парная, узкая мышца, начинающаяся от седалищных бугров. Она направляется к срединной линии, где соединяется с одноимённой мышцей противоположной стороны, участвуя в образовании сухожильного центра промежности.
Сфинктер мочеиспускательного канала (m.sphincter urethrae) – парная мышца, лежит кпереди от предыдущей. Периферически расположенные пучки этой мышцы направляются к ветвям лобковых костей и к фасциям мочеполовой диафрагмы. Пучки этой мышцы окружают мочеиспускательный канал. Эта мышца соединяется с влагалищем.
РАЗДЕЛ 7
ПОЛОВОЕ ВОСПИТАНИЕ КАК ОСНОВА ФОРМИРОВАНИЯ ЗДОРОВОЙ СЕМЬИ И ПРОФИЛАКТИКИ БОЛЕЗНЕЙ, ПЕРЕДАЮЩИХСЯ ПОЛОВЫМ ПУТЕМ
7.1. Анатомия и физиология репродуктивной системы
Репродуктивная система состоит из мужских и женских половых органов, но они сами, без соответствующего физического и репродуктивного здоровья, не способны обеспечить продолжение человеческого рода. Репродуктивное здоровье дает возможность людям иметь полноценных потомков. Репродуктивная функция проявляется лишь в определенном возрасте, который является весьма ограниченным, особенно с учетом современных экологических обстоятельств, способных влиять на состояние и функцию половых органов и вызывать мутации в геноме всех клеток. Только физически, духовно и репродуктивно здоровые люди могут иметь жизнеспособных, полноценных потомков.
Концепция ВОЗ по здоровому половой жизни каждого человека основывается на таких принципах:
На возможности получать наслаждение от половых отношений, иметь детей, контролировать свое поведение согласно требованиям общества и личной этики;
На свободе от страха и стыда, чувство вины, предрассудков и других психологических факторов, которые мешают половым отношениям;
Отсутствие органических расстройств, болезней и недостатков, которые ограничивают половые и репродуктивные функции.
Внутренние органы половой системы человека формируются на 10-12 неделю внутриутробной жизни под влиянием генетических факторов, поэтому сначала они имеют одинаковую структуру независимо от пола. С 12 недели их дальнейшее развитие зависит от образования мужских половых гормонов, которые обеспечивают развитие внешних органов мужчин, тогда как у эмбрионов, не имеющих этих гормонов, формируются женские наружные половые органы.
Половая система мужчины состоит из парных половых желез (семенники, тестикулы или яички), семявыносящих протоков, придаточних желез (міхурцеві, предстательная, бульбоуретральні) и полового члена (пенис или фаллос).
Половые железы находятся в мошне, в них образуются спермии (половые клетки или гаметы) и половые гормоны (стероидные вещества, которые обеспечивают развитие первичных и вторичных половых признаков, образование и созревание спермиев, половую потенцию).
Семенники имеют размер 3x5 см, они построены из почти тысячи извилистых канальцев, диаметром по 200 мкм, а их общая длина 300-400 м. Стенки извитых семенных канальцев у половозрелого мужчины содержат сперматогенні клетки - сперматогонии, из которых постоянно образуются мужские половые клетки от времени полового созревания и до климактерического периода.
В канальцах между половыми клетками, которые находятся на разных этапах развития, расположены клетки Сертоли. Они содержат много органелл, которые обеспечивают образование питательных веществ для сперматогоній и жидкость, в составе которой спермии проходят канальцами.
Пространство между канальцами заполняют клетки Лейдига, в которых образуются половые гормоны - (андрогены - от andros - «мужчина»), прежде всего тестостерон, они влияют на секреторные клетки придаточных половых желез, кожу, половое развитие и половое созревание, половое поведение (либидо и потенцию), стимулируют синтез белков, ускоряют рост тканей, но в определенном возрасте тормозят рост трубчатых костей в длину. При низкой концентрации гормонов сперматогенез растет, а при высокой - уменьшается. В семенниках синтезируются в небольшом количестве еще и женские половые гормоны. Во время старения уменьшается количество половых клеток - сперматогоній, но высокая сексуальная активность задерживает и тормозит этот процесс.
Между кровью и тканями семенников существует барьер, который называется гемотестикулярним, он способствует сохранению сперматогоній, потому что не пропускает из крови токсические вещества, лейкоциты и антитела.
До заднего края каждого сім"яника плотно прилегает его придаток, длиной 5-6 см, шириной 0,5-1 см. Придаток является системой с 10-20 отдельных канальцев общей длиной 6-8 м. Они заполнены сперматозоидами, которые приобретают здесь способности к оплодотворению и отрицательный заряд на своей поверхности.
Семявыносящие протоки - отходят от каждого придатка сім"яника, проходят через подмышечный канал, а возле мочевого пузыря оба протока сливаются. Длина каждого пролив равна 30-40 см, внешний диаметр - 3-5 мм, а внутренний просвет - 1-2 мм. Конечный отдел протока образует ампулу. Далее сперма поступает в сім"явипорскувальну пролив длиной 2 см, которая проходит через предстательную железу и открывается в простатну часть мочеиспускательного канала (уретры), что проходит в половом члене.
Семь яли пузырьки - придаточные половые железы, которые находятся рядом с ампулами семявыносящих протоков. Это парные образования размером 5x2x1 см, которые выделяют секрет желтого цвета, густой по консистенции, он смешивается со спермой, выполняет трофическую роль, активизирует сперматозоиды.
Простата (предстательная железа) - это железисто-мускульный орган массой 18-22 грамм. Она окружает начальную часть мочеиспускательного канала, к которому открываются ее протоки. Через них выделяется жидкий беловатый секрет с простагландинами и другими биологически активными веществами. Этот очень важный орган часто называют «вторым сердцем мужчины».
Бульбоуретральні (Куперові) железы-маленькие образования, их протоки открываются в просвет мочеиспускательного канала. Железы выделяют секрет, что защищает слизистую мочеиспускательного канала от раздражающего действия компонентов мочи.
Половой член (penis, phallos) - образование, которое обеспечивает поступление спермы к шейке матки. Половой член построен из двух пещеристых тел и одного губчатого тела, которое формирует его головку. Задняя часть члена образует луковицу, размещенную в глубине мышц промежности.
Во всех древних культурах фаллическая символика играла большую роль. По Фрейду, мальчики считают, что все люди, в том числе женщины, имеющие пенис, поэтому у них возникает страх его потери, а девочки хотят его иметь.
У народов, исповедующих ислам и иудаизм, обрезание (удаление крайней плоти) является важным религиозным обрядом, который символизирует вечный завет между богом и людьми (Бытие,7:9-14). Они считают, что крайняя плоть является женской по своей сути, потому что в ней скрывается головка полового члена.
Сперма - густая, беловатая или желтоватая жидкость с запахом свежих каштанов. Она образуется смешиванием секретов половых и придаточних желез, благодаря которым жидкость из семенников разбавляется, ее объем значительно возрастает, повышаются жизнеспособность и активность сперматозоидов. В состав спермы входит вода, слизь, фруктоза, лимонная кислота, простагландины, которые вызывают сокращение гладких мышц влагалища, матки, маточных труб.
У здорового взрослого мужчины в 1 мл спермы помещается примерно 40-120 млн сперматозоидов, а во время одной еяколяції (извержение спермы) выделяется до 300-400 млн в общем объеме 2-5 миллилитр.
Сперматозоид человека имеет головку, шейку и хвостик. Головка содержит ядро с 23 хромосомами, в том числе одной половому (х или у), а шейка и хвостик - митохондрии и микротрубки, благодаря которым спермий способен к самостоятельному перемещению. Процесс образования каждого
сперматозоида составляет 70 суток, ежесуточно у мужчины образуется их более 100 млн.
Развитие и функция половых желез зависит от гонадотропных гормонов гипофиза. Фолликулостимулирующий гормон обеспечивает развитие клеток Сертоли и спермы, лютеинизирующий - синтез андрогенов, пролактин - рост простаты и семенных пузырьков. Под влиянием андрогенов происходит развитие вторичных половых признаков - рост наружных половых органов, функционирования придаточних желез, увеличение мышечной ткани, низкий голос, выраженность и локализация волос на теле, особенности полового поведения.
Половая система у женщин состоит из внутренних и внешних органов. К внутренним относятся - парные яичники, яйцеводы (маточные или Фалопиевые трубы), матка, влагалище (вагина). К внешним относят вульву (большие и малые половые губы, окружающие вход во влагалище) и клитор.
Яичники - парные железы смешанной секреции, расположенные в брюшной полости, в них созревают половые клетки и образуются половые гормоны - эстрогены фолікулярними клетками (от oestrus - течка), преимущественно эстрадиол и гестагены клетками желтого тела (от hystera-матка), а именно прогестерон. Вес яичника у молодой взрослой девушки (женщины) 5-6 г, в возрасте 40-50 лет он начинает атрофироваться. В бархатном слое находятся фолликулы, которые развиваются, а в мозговом слое - сосуды и зрелые фолликулы.
Формируется яичник в эмбриональный период, когда первичные половые клетки превращаются в овогонії (от ovum - яйцо). В это время происходит размножение женских половых клеток митозом, образуются первичные фолликулы с овоцитами первого порядка. У новорожденной девочки в обоих яичниках находится до 2 млн женских половых клеток, часть из них в дальнейшем может размножаться, а часть погибает. Ко времени полового созревания остается почти 300000 первичных овоцитів. Дальнейшие изменения происходят после полового созревания, когда начинаются менструальные циклы, повторяющиеся через каждые 28 суток и завершается овуляцией.
Яйцеклетка - это женская половая клетка, развивающаяся в яичнике женщины. Имеет округлую форму, неподвижная, величина - 130-140 мкм. Яйцеклетка содержит ядро, цитоплазму и оболочки. В ядре яйцеклетки содержится гаплоидный набор хромосом: 22 автосоми и 1 половая Х-хромосома.
Процесс развития женских половых клеток называется овогенезу. Различают три периода овогенезу: размножения, роста и созревания. Период размножения происходит во время эмбрионального развития. Период созревания завершается после овуляции.
Яйцепроводы - мышечные трубчатые образования длиной по 12 см, которыми половые клетки попадают от яичников до матки.
Матка - мышечный мешок, который внутри содержит много железистых клеток, его объем во время беременности увеличивается с 10 см3 до 5000 см3. Матка соединяется с влагалищем через шейку матки.
Влагалище - эластичная мышечная трубка в области малого таза, через которую матка соединяется с внешними половыми органами.
Вульва - половые губы, то есть складки кожи (большие) и слизистые (малые), которые закрывают вход во влагалище. Содержит бартолінієві железы, которые при половом возбуждении декретують слизь увлажняет влагалище. В месте соединения малых половых губ находится клитор - аналог полового члена мужчин.
Женские половые клетки начинают развиваться в период полового созревания (13-16 лет). При половом созревании у девочек возникают периодические колебания количества эстрогенов в крови и моче, соответственно периодических колебаний деятельности половых желез, которые обеспечивают женский половой цикл. Во время одного из очередных повышений уровня эстрогенов проходит овуляция - разрыв одного из яичниковых фолликулов и выход наиболее зрелой яйцеклетки. Через несколько дней после этого начинается первая менструация. У девочек половые циклы нерегулярные. Половая зрелость наступает в полной мере лишь тогда, когда циклы становятся регулярными и овуляторними.
Половой цикл женщины в среднем длится 27-28 дней. За этот период созревает фолликул, а в конце срока происходит овуляция. Яйцеклетка по маточным трубам передвигается в матку. Если произошло оплодотворение, яйцеклетка закрепляется в стенке матки, если же не произошло - удаляется из организма.
На месте фолликула образуется желтое тело, которое продуцирует гормон - прогестерон, который противодействует стимулирующему влиянию эстрогенов на сокращение мышц матки и труб. При оплодотворении яйцеклетки и наступлении беременности желтое тело сохраняется и разрастается, если же оплодотворения не произошло - исчезает. Исчезновение желтого тела создает условия для созревание следующего фолликула.
С периодическим созреванием фолликула и овуляцией связано истечение крови из матки - менструация. Она наступает циклически через каждые 27-28 дней, если оплодотворения не произошло. Менструация (менархе) связана с изменениями в слизистой оболочке матки. Последняя набухает и разрастается, готовясь к принятию оплодотворенной яйцеклетки; если оплодотворение не произошло, слизистая оболочка разрушается и удаляется из матки. Удаление сопровождается вытеканием крови. Менструация длится 3-5 дней, при этом теряется от 50 до 250 мл крови. Наступления менструации происходит благодаря уменьшению в этот период концентрации в крови гормонов яичников, что приводит к тонических сокращений матки. После менструации слизистая оболочка регенерируется. Весь цикл снова повторяется.
В возрасте после 50-ти лет у женщин начинается климактерический период. который характеризуется потерей половых функций. Половые циклы становятся нерегулярными, безовуляторними, наступает аменорея - прекращение менструации.
У мужчин климакс наступает после 60-ти лет и характеризуется потерей подвижности сперматозоидов, утратой способности к оплодотворению, к эякуляции - семяизвержения.
Наступление климакса индивидуальное и зависит от образа жизни, перенесенных заболеваний, климата, питания.
Для правильной клинической оценки нейроэндокринных нарушений в организме женщины и, соответственно, определения принципов и мето-дов их патогенетической терапии необходимо прежде всего знание пятизве-ньевой регуляции репродуктивной системы, основной функцией которой является воспроизводство биологического вида (рис. 1).
Регуляция функции репродуктивной системы определяется гипоталамо-гипофизарным звеном, которое, в свою очередь, посредством нейромедиато-ров и нейротрансмиттеров контролируется корой головного мозга (Lakoski J.M., 1989).
Гипоталамус является своеобразными биологическими часами организма, то есть системой саморегулирования и автоматизации нейрорегуляторных процессов, которая реализует информацию, поступающую из внешней и вну-тренней среды организма, обеспечивая тем самым внутренний гомеостаз, не-обходимый для нормального течения физиологических процессов. Именно гипоталамус является тем ключевым звеном, координирующим деятельность гипоталамо-гипофизарно-яичникового комплекса, функция которого регули-руется как нейропептидами ЦНС, так и яичниковыми стероидами по механиз-му обратной связи (Wildt L., 1989; Sopelak V.M., 1997).
Учитывая достаточно хорошую освещённость в современной литературе периферического звена репродуктивной системы, а также увеличение роли всё возрастающих психоэмоциональных нагрузок в Рисунок 1. Функциональная структура репродуктивной системы механизмах развития дис-гормональных нарушений, мы сочли целесообразным более подробно остано-виться на некоторых аспектах участия надгипоталамических структур в ре-гуляции репродуктивной системы.
Как известно, мозг состоит из двух типов клеток: из нейронов, составляю-щих 10% всех клеток мозга и из глии — астроцитов и олигодендритов, состав-ляющих, соответственно, остальные 90%.
Развитие нейронов и глии происходит из нейроэпителиального предшест-венника — стволовой клетки, в результате развития которой происходит синтез 2-х клеточных линий: нейрональных клеток-предшественников, из которых возникают различные типы нейронов, и глиальных клеток-предшест-венников, из которых в дальнейшем развиваются астроциты и олигодендро-циты (Lakoski J.M., 1989; Sopelak V.M., 1997).
Нейроны — это высокодифференцированные клетки с четкими размерами, формой и внутриклеточными органеллами. Как и все другие клетки, за исклю-чением эритроцитов, нейроны имеют тело клетки, в центре которого распола-гается ядро, окруженное различным объемом цитоплазмы.
От поверхности нейронов ответвляются воспринимающие отростки — дендриты и единственный главный передающий отросток — аксон, который простирается к своим специфическим синаптическим клеткам-мишеням и мо-жет значительно варьировать по длине (Sopelak V.M., 1997).
Ключевой процесс жизнедеятельности нейрона концентрируется в цито-плазме тела клетки (она также называется перикарионом), и затем продукты нейронального синтеза транспортируются в аксоны и дендриты. Двухсторон-ний транспорт между участками тела клетки и дистальными отростками обес-печивает целостность нейронной функции и является постоянным энергети-чески-зависимым слаженным процессом.
Клетки глии (от английского слова glue — клей) первоначально рассматри-вали как поддерживающие клетки мозга, но исследования последних лет опре-делили их важную функциональную роль в регуляции жизнедеятельности ней-ронов. Этот класс ненейронных клеточных элементов, в 9 раз превышающий количество нейронов, фактически обеспечивает взаимодействие между ними.
Наиболее многочисленные глиальные клетки названы астроцитами, бла-годаря их мультиотростковым очертаниям. Эти клетки характеризуются уни-кальной экспрессией глиального фибриллярного кислотного протеина и рас-положены между наружной поверхностью сосудов, нейронами и их соедине-ниями (рис. 2). Отростки астроцитов направляются от нейронов к капилля-рам, где они формируют периваскулярное основание.
 Капиллярное основание астроцитов охватывает около 85% капилляров че-ловеческого мозга и формирует гемато-энцефалический барьер.
Капиллярное основание астроцитов охватывает около 85% капилляров че-ловеческого мозга и формирует гемато-энцефалический барьер.
Другой ваный класс клеток глии — олигодендроциты (клетки с малым количеством коротких и толстых отростков), которые формируют миелино-вую оболочку аксонов, что позволяет нейронам осуществлять свой эффект быстро и без ослабления на длинных расстояниях в пределах нервной систе-мы. Олигодендроциты также содержат ферменты стероидного генеза Р450 и продуцируют прегнанолон из холестерола.
Определение в ткани мозга ферментов стероидогенеза явилось одним из открытий, способствующим раскрытию механизмов участия ЦНС в регуля-ции репродуктивной функции и, что не менее важно, объясняющих измене-ния в ЦНС под влиянием изменений гормонального гомеостаза.
Секреция нейроактивных стероидов в астроцитах выше, чем в олигоденд-роцитах и нейронах, в связи с чем следует более детально остановиться на ха-рактеристике именно этих клеток.
Свойства астроцитов различны и ещё до конца не изучены, хотя уже сей-час существуют доказательства того, что астроциты являются паракринными клетками для нейронов:
В астроцитах выявлено наличие инсулиноподобного фактора роста (ИФР), содержание которого увеличивается к периоду полового созревания, а также растёт при лечении эстрогенами;
Питуициты, как тип астроцитов, являются главными ненейронными кле-точными элементами в нейрогипофизе и играют важную роль в контроле вы-броса окситоцина и вазопрессина из нейросекреторных нервных окончаний;
Присутствие в астроцитах рецепторов лютеинизирующего гормона (ЛГ) и человеческого хорионического гонадотропина (ХГ) предполагает, что ЛГ и ХГ могут влиять на функцию глиальных клеток и, соответственно, на процес-сы развития и функционирование мозга;
Астроциты способны продуцировать множество иммуномодулирующих молекул, таких как интерлейкины (ИЛ-1, ИЛ-2, ИЛ-6), туморнекрозный фактор а, трансформирующий фактор роста-ос, интерферон и простагландин Е, при этом пролактин индуцирует митогенез и экспрессию цитокинов в астроцитах;
Астроциты, как и нейроны, способны продуцировать кортикотропин-ри-лизинг фактор связывающий протеин (КРФ-СП), широко представленный в мозге. Стероиды, такие как дексаметазон, гидрокортизон и, в меньшей степе-ни, дегидроэпиандростерон, угнетают выброс КРФ-СП из астроцитов;
Астроциты гипоталамического происхождения секретируют трансфор-мирующий фактор роста а и (3, который стимулирует генную экспрессию го-надотропин-рилизинг гормонов (Гн-РГ) в нейронах, при этом гипоталамичес-кие астроциты примерно в 4 раза активнее, чем астроциты коры в отношении синтеза дегидроэпиандростерона (ДГЭА).
Астроциты также могут участвовать в регуляции нейротрансмиттерного уровня глутамата, обеспечивающего возбуждающий эффект, и у-аминомасля-ной кислоты (ГАМК), играющей ключевую роль в достижении анксиолитиче-ского (успокаивающего) эффекта.
В настоящее время выделены 3 главные химические формы трансмитте-ров: аминокислоты, моноамины и нейропептиды.
Аминокислоты действуют в качестве трансмиттеров как возбуждающе, так и угнетающе. В возбуждающих соединениях трансмиттерных субстанций ключевыми являются ацетилхолин, а также глутамат и аспартат. Ингибитор-ные соединения регулируются такими аминокислотами, как ГАМК и глицин.
Моноамины, как трансляторы, состоят из катехоламинергических (адрена-лин, норадреналин и допамин) и серотонинергических трансмиттеров. Так, тирозин поступает из кровотока внутрь катехоламиновых нейронов и являет-ся субстратом, из которого тирозин-гидроксилаза катализирует синтез допа. Трансформация допа в допамин происходит с помощью аминокислоты декар-токсилазы (АКД). Допамин-(3 оксидаза (ДВО) в норадренергических нейро-нах трасформирует допамин в норадреналин (НА).
ДА и НА высвобождаются в синаптическую щель, где они быстро связы-ваются с постсинаптическими рецепторами. В плазме избыток трансмиттеров претерпевает либо метаболическую инактивацию с помощью катехол-О-ме-тилтрансферазы (КОМТ), либо обратный захват пресинаптическими рецепто-рами, где они претерпевают метаболическую деградацию с помощью моно-аминооксидазы (МАО), формируя дегидроксифенилэтилгликоль (ДОФЭГ).
Пептидные трасмиттеры. Пептид-содержащие нейроны гипоталамуса были первоначально описаны как нейросекреторные нейроны, но позже стало известно, что практически все гипоталамические нейропептиды проецируют-ся во многие области мозга. Они обеспечивают нейротрансмиттерные функ-ции в регуляции приема пищи, пищевого и сексуального поведения (табл. 1).
Отдельно следует остановиться на роли оксида азота в центральной и пе-риферической нервной системе, открытие которого радикально изменило су-ществовавшие ранее взгляды на синаптическую трансмиссию. Хотя имеются существенные доказательства того, что оксид азота функционирует как ней-ротрансмиттер, следует отметить, что это необычный трансмиттер, т.к. он яв-ляется лабильным газом, который не может храниться в синаптических пу-зырьках. Оксид азота синтезируется из L-аргинина с помощью оксидазот-синтетазы и из нервных окончаний попадает путем простой диффузии, а не путем экзоцитоза как остальные нейротрансмиттеры (рис. 3). Более того, ок-сид азота не претерпевает обратимых реакций с рецепторами, как все осталь-ные обратимые нейротрансмиттеры, а формирует ковалентные соединения с несколькими потенциальными мишенями, которые включают ферменты, та-кие как изонилатциклаза и другие молекулы.
: Действие обратимых нейротрансмиттеров ограничено пресинаптическим выбросом или ферментной деградацией, в то время как действие оксида азо-та обеспечивается диффузией вдали от мишеней или формированием кова-лентных соединений с супероксидным анионом.
Формирование оксида азота из аргинина в мозге катализируется с помо-щью оксидазот-синтетазы в присутствии кислорода с НАДФ как кофермента и тетрагидробиопротеина как кофактора.
Таблица 1 Пептидные трансмиттеры в ЦНС
(по Yen S.S.C., 1999 с изменениями и дополнениями)
|
Гипофизотропные гормоны |
Гастроинтестинальные I пептиды |
|
Тиротропин-рилизинг гормон |
Вазоактивный интестинальный полипептид |
|
Гонадотропин-рилизинг гормон |
Холицистокинин |
|
Кортикотропин-рилизинг гормон и урокортин |
Гастрин |
|
Соматотропин-рилизинг гормон |
Субстанция П |
|
Соматостатин |
Нейротензин |
|
Нейрогипофизарные гормоны |
Метионин энкефалин |
|
1Вазопрессин |
Лейцин энкефалин |
|
Окситоцин |
Динорфин |
|
Нейрофизин 1 и N |
Неоэндорфин |
|
Другие |
Инсулин |
|
Нейропептид Y |
Глюкагон |
|
Ангиотензин i |
Бомбезин |
|
Брадикинин |
Секретин |
|
Карнозин |
Соматостатин |
|
Кальцитонин ген-зависимый пептид |
Мотилин |
|
Интерлейкин-1 |
Гипофизарные пептиды |
|
Ингибин |
Адренокортикотропин |
|
Панкреастатин |
р-эндорфин |
|
Амилин |
а-меланоцит стимулирующий гормон |
|
Паратгормон-зависимый пептид |
Пролактин |
|
Активин |
Лютеинизирующий гормон |
|
Гормон роста Тиротропин |
Относительно роли оксида азота в центральной регуляции репродуктивной системы, следует отметить, что N0 является нейротрансмиттером, регулирующим выброс Гн-РГ.
Нейростероиды.
Открытие в гипоталамусе локального синтеза эстрогенов (Naftollin et al, 1975) позволило предположить, что мозгу свойственна функ-ция стероидогенеза. В 1981 г. в мозге взрослых крыс-самцов обнаружено нали-чие прегнанолона и прегнанолона-сульфата, а также дегидроэпиандростерона (ДГЭА) и дегидроэпиандростерона сульфата (ДГЭА-С). Это привело к откры-тию механизмов биосинтеза стероидов в ЦНС, названных нейростероидами.
В человеческом мозге нейростероиды, как и нейротрансмиттеры, обнару-жены у мужчин и у женщин старше 60 лет. ДГЭА, прегнанолон и прогестерон присутствуют во всех участках мозга, при этом их концентрация в мозге в не-сколько раз выше, чем в плазме.
В мозге выявлено также наличие ДГЭА-сульфаттрансферазы и сульфа-тазы, следовательно можно предположить, что синтез ДГЭА-С происходит непосредственно в мозге.
Стероидогенный фактор-1 (СФ-1) — тканеспецифический ядерный рецептор — регулирует гены нескольких ферментов стероидогенеза и широко представлен в человеческом мозге, включая компоненты лимбической системы.
Нейростероиды играют чрезвычайно важную роль во всех процессах жиз-недеятельности организма, они модулируют активность ГАМК-рецепторов, глутамат-рецепторов, влияют на когнитивную функцию, оказывают трофиче-ское действие на нервную ткань (способствуют миелинизации), модулируют выработку рилизинг-гормонов в гипоталамусе (Yen S.S.C., 1999).
Гипоталамус — это часть диэнцефалон, лежащая под третьим желудоч-ком между зрительным перекрестом и срединным возвышением, которая соединяется с задней долей гипофиза посредством гипофизарного ствола, а также соединяется с парными сосцевидными телами (рис. 4). Гипоталамус взаимосвязан с ЦНС и гипофизом посредством множества циркуляторных и нейронных связей. Он состоит из нервных клеток, сгруппи-рованных в ядра. Клетки, сгруппированные в паравентрикулярные и супраоп-тические ядра гипоталамуса, продолжаются до задней доли гипофиза, где происходит высвобождение вазопрессина, окситоцина и нейрофизинов. При этом супраоптические и паравентрикулярные ядра имеют прямую нейронную связь с задней долей гипофиза. Супраоптические ядра секретируют главным образом вазопрессин, а паравентрикулярные — окситоцин, который транс-портируется по нервным окончаниям в заднюю долю (Sopelak V.M., 1997).
Рисунок З. Формирование оксида азота в мозге (Yen S.S.C., 1999)
Другие ядра продуцируют рилизинг- и ингибиторные факторы (Гн-РГ, ТРГ, соматостатин, кортикотропин-рилизинг гормон (КРГ), которые транс-портируются в переднюю долю гипофиза по кровеносной портальной систе-ме и контролируют секрецию передней доли гипофиза.
Функциональные связи с передней долей гипофиза представлены системой гипоталамо-гипофизарных кровеносных сосудов (Wildt L., 1989). Гипоталами-ческие гормоны попадают в переднюю долю через медиальное возвышение и гипоталамо-портальный кровоток. Гипоталамус также имеет интрагипотала-мические нейронные соединения, афферентные волоконные соединения со средним мозгом и лимбической системой, эфферентные волоконные соедине-ния со средним мозгом и лимбической системой, а также с задней долей гипо-физа. Гипоталамические факторы транспортируются по нервным волокнам в срединное возвышение, где они проникают через стенки капилляров гипофиза (рис. 5). Эти факторы влияют на эндокринные клетки гипофиза и обеспечива-ют специфические гормональные ответы (Yen S.S.C., 1999).
Говоря о регуляции репродуктивной системы, следует подчеркнуть, что под влиянием рилизинг-гормонов гипоталамуса осуществляется синтез гонадотропных гормонов в гипофизе. Местом синтеза гипофизотропных рилизинг-гормонов (либеринов), представляющих по химической природе декапептиды, являются именно аркуатные ядра медиобазального гипоталамуса. Выработка рилизинг-гормонов происходит в определенном пульсирующем ритме, названном цирхоральным.
Для обеспечения нормальной секреции гонадотропинов достаточно под-держания стабильной частоты выброса физиологических количеств Гн-РГ. Из-менение частоты выброса Гн-РГ меняет не только количество ЛГ и ФСГ, вы-деляемых гипофизом, но и их соотношение, в то время как даже десятикратное повышение концентрации Гн-РГ ведет только к небольшому повышению выделения ФСГ и никак не меняет секреции ЛГ (Halvorson L.M. et al., 1999).
Так, повышение ритма ведет к значительному повышению выброса ФСГ и к снижению выброса ЛГ. В лютеиновую фазу прогестерон через эндогенные опиаты урежает частоту пульсового генератора, причем данное действие оп-ределяется не концентрацией прогестерона, а длительностью его воздействия. Эстрадиол, действуя на гипоталамус и на гонадотропы (увеличение плотнос-ти рецепторов Гн-РГ), повышает амплитуду волны ЛГ/ФСГ.
|
|
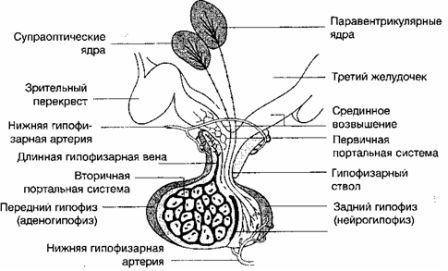
Частота выброса Гн-РГ у человека составляет 1 выброс в 70-90 минут и соот-ветствует целому ряду биоритмов (чередованию фаз сна, колебанию скорости клубочковой фильтрации и желудочной секреции, частоте приливов во время климакса и т.д). Частотная модуляция информации обеспечивает быстроту и на-дежность регуляции репродуктивной системы и ее устойчивость к помехам.
Пульсовой генератор ритма — аркуатное ядро гипоталамуса в физиологи-ческих условиях получает информацию о выделении гонадотропинов гипофи-зом по системе короткой обратной связи, так как специальные сфинктеры ре-гулируют градиенты давлений в воротной системе кровотока, и часть крови из гипофиза поступает обратно в гипоталамус, что обеспечивает очень высокую местную концентрацию гормонов гипофиза в гипоталамусе (Yen S., 1999).
Синтез и секреция ЛГ и ФСГ в гипофизе осуществляются одними и теми же клетками (Halvorson L.M. et al., 1999). На поверхности гонадотрофов име-ются рецепторы к Гн-РГ, плотность которых зависит от уровня стероидных гормонов в крови и от концентрации Гн-РГ. Соединение Гн-РГ с рецептором вызывает массивное поступление ионов кальция внутрь клетки, что через не-сколько минут ведет к выбросу запаса ЛГ и ФСГ в кровоток. Кроме того, Гн-РГ вызывает стимуляцию синтеза ЛГ и ФСГ и поддерживает целостность гонадотрофов (Wildt L., 1989).
Важная роль в регуляции функции желез внутренней секреции принадле-жит гипофизу. Он лежит в турецком седле в основании мозга, состоит из пе-редней (аденогипофиз), промежуточной и задней (нейрогипофиз) долей. Про-межуточная доля у человека практически отсутствует. Гипофиз соединяется с гипоталамусом через гипофизарный ствол (см. рис. 5).
Передняя доля гипофиза складывается из пяти различных типов клеток, различающихся по иммунологическим и ультраструктурным характеристи-кам. Эти клетки в передней доле продуцируют 6 известных гормонов:
Адренокортикотропный гормон (АКТГ), или кортикотропин;
Тиреотропный гормон (ТТГ), или тиреотропин;
Гонадотропные гормоны: фолликулостимулирующий (ФСГ), или фолли-тропин, и лютеинизирующий (ЛГ), или лютропин;
Соматотропный гормон (СТГ), или гормон роста;
Пролактин.
Первые 4 гормона регулируют функции так называемых периферических желез внутренней секреции, а соматотропин и пролактин действуют непо-средственно на ткани-мишени (Halvorson L.M. et al., 1999).
Гормон роста и пролактин продуцируются двумя типами клеток — соматотрофами и лактотрофами (маммотрофы), принадлежащими к ацидо-фильной серии. АКТГ и другие фракции молекул проопиомелатокортина, та-кие как р-липотропин и эндорфины синтезируют тиротрофами, а ЛГ и ФСГ — гонадотрофами, принадлежащими к базофильной серии.
Гонадотрофы составляют 10-15% от клеточного состава передней доли ги-пофиза и располагаются вблизи лактотрофов. Эта особенность локализации позволяет предположить, что между двумя видами этих клеток имеются пара-кринные взаимоотношения (Sopelak V.M., 1997).
Как уже упоминалось, секреция этих шести гормонов передней доли кон-тролируется гипоталамическими рилизинговыми и ингибиторными фактора-ми, которые секретируются в гипоталамусе и попадают в гипофиз через гипо-таламо-гипофизарные портальные сосуды. Однако на продукцию тропных гормонов могут влиять и другие субстанции, синтезируемые как в централь-ных (Р-эндорфины), так и в периферических (эстрадиол) отделах репродук-тивной системы (Halvorson L.M. et al., 1999).
Нейрогипофиз включает гипофизарный ствол (см. рис. 5), нейральную до-лю и срединное возвышение (специальная невральная ткань в основании ги-поталамуса, формирующая главный регион для переноса гипофиз-регулирую-щих нейросекретов в переднюю долю гипофиза). Два гормона задней доли гипофиза (вазопрессин и окситоцин) накапливаются в гранулах с соответст-вующими нейрофизинами, транспортируются по аксонам и собираются в тер-минальных отделах аксонов, где они хранятся до соответствующих импуль-сов, которые вызывают их выброс. Нейропептиды освобождаются из секре-торных гранул путем экзоцитоза. Этот процесс включает растворение мемб-ран нейросекреторных гранул и небольшого участка клеточной мембраны на окончании аксона. Содержимое гранул попадает в межклеточное пространст-во, а оттуда — в кровоток (Sopelak V.M., 1997) .
Регуляция репродукции и функция гонад осуществляются преимуществен-но гонадотропными гормонами, секретируемыми аденогипофизом, а именно ФСГ, ЛГ и пролактином. ФСГ — вызывает пролиферацию гранулезных клеток, стимулирует рост фолликулов. ЛГ — активизирует синтез андрогенов и совместно с ФСГ способствует овуляции. Секреция ФСГ и ЛС регулируется гонадотропин-рилизинг гормоном по механизму обратной связи и зависит так-же от уровня эстрогенов и андрогенов. Гонадолиберин (люлиберин) секрети-руется пульсами с частотой от 1 пульса в час до 1-2 пульсов за сутки. Контроль секреции гонадолиберина осуществляется половыми и другими гормонами, многочисленными нейротрансмиттерами ЦНС, включая катехоламины, опиат-ные гормоны и др. Гонадолиберин взаимодействует с рецепторами, располо-женными на мембранах гонадотрофов, и для активации рецептора требуется обязательное наличие трех первых аминокислот. Агонисты гонадолиберина (бузерилин, нафарелин, леупролид и др.) оказывают свой эффект посредством взаимодействия с теми же мембранными рецепторами (Halvorson L.M., 1999).
Пролактин угнетают продукцию гонадотропных гормонов. Угнетающее действие на выделение ЛГ оказывают также глюкокортикоиды.
По химической структуре ЛГ и ФСГ — гликопротеиды, состоящие из двух полипептидных субъединиц а и р. а-субъединица этих гормонов является об-щей для каждого гликопротеида и имеет одинаковую последовательность ами-нокислот, Р-субъединица отличается среди гликопротеидов по последователь-ности расположения входящих в неё аминокислот. Именно Р-субъединица от-вечает за гормональную специфичность. Обе субъединицы по отдельности би-ологически неактивны. Образование гетеродимеров является обязательным условием для проявления биологической активности (Halvorson L.M., 1999).
Период полураспада гонадотропинов, циркулирующих в крови, имеет пря-мое отношение к компоненту сиаловых кислот в молекуле гормона. Доказано, что десиалирование укорачивает период полураспада и биологическую актив-ность гонадотропинов. ФСГ находится в крови в свободной форме и период его полураспада составляет 55-60 мин, а ЛГ — 25-30 мин. В репродуктивном возрасте ежедневное высвобождение ЛГ составляет 500-1100 мМЕ, в постме-нопаузе скорость образования ЛГ увеличивается и его количество составляет до 3000-3500 мМЕ в день (Sopelak V.M., 1997).
Подобно стероидам, гонадотропины оказывают биологический эффект на ткани-мишени посредством активации специфических рецепторов. Однако в отличие от стероидных гормонов, рецепторы гонадотропинов связаны с мем-браной клеток-мишеней. Поверхностные клеточные рецепторы к пептидным гликопротеиновым гормонам являются белками, входящими в структуру кле-точной мембраны. После связывания с гонадотропином мембранные рецепто-ры стимулируют выработку растворимых внутриклеточных мессенджеров, которые, в свою очередь, обеспечивают клеточный ответ (Halvorson L.M., ChinW.W., 1999).
Регуляторами выработки ФСГ, согласно современным представлениям, помимо гипоталамических либеринов являются ингибин и активин, которые продуцируются гранулёзными и лютеиновыми клетками яичников, а также клетками цитотрофобласта (Hopko Ireland et al, 1994).
Ингибин состоит из двух субъединиц аир. ФСГ влияет на синтез и выде-ление ингибина по принципу обратной связи. Комбинация ос-субъединицы с (3-субъединицей приводит к супрессии ФСГ, а комбинация двух (3-субъеди-ниц приводит к образованию активина и, таким образом, к стимуляции ФСГ.
Влияние на синтез и выделение ФСГ оказывает также фоллистатин, выде-ленный из фолликулярной жидкости. Фоллистатин является гликопротеидом, который подобно ингибину уменьшает высвобождение ФСГ в культуре гона-дотропных клеток гипофиза. Кроме того, он имеет высокую аффинность к связыванию активина и менее выраженную к связыванию ингибина. Установ-лено, что фоллистатин и активин А являются компонентами аутокринно-па-ракринной системы фолликула и участвуют в регуляции различных функций клеток внутренней оболочки граафова пузырька (Grome N., O"Brien ML, 1996).
Существуют 3 типа секреции гонадотропинов: тонический, циклический и эпизодический, или пульсирующий (Halvorson L.M., Chin W.W., 1999).
Тоническая, или базальная, секреция гонадотропинов регулируется по-средством отрицательной обратной связи, а циклическая — механизмом по-ложительной обратной связи с участием эстрогенов.
Пульсирующая секреция обусловлена активностью гипоталамуса и высво-бождением гонадолиберинов.
Развитие фолликула в первой половине цикла осуществляется благодаря тонической секреции ФСГ и ЛГ. Повышение секреции эстрадиола приводит к торможению образования ФСГ. Развитие фолликула зависит от количества рецепторов к ФСГ в клетках гранулезной зоны, причем синтез этих рецепто-ров, в свою очередь, стимулируется эстрогенами.
Та^им образом, ФСГ приводит к синтезу в определенном фолликуле эст-рогенов, которые, увеличивая количество рецепторов к ФСГ, способствуют его накоплению (путем связывания его рецепторами), дальнейшему созрева-нию фолликула и увеличению секреции эстрадиола. Другие фолликулы в это время подвергаются атрезии. Концентрация эстрадиола в крови достигает максимума в предовуляторный период, что приводит к высвобождению боль-шого количества гонадолиберина и последующего пика высвобождения ЛГ и ФСГ. Предовуляторное повышение ЛГ и ФСГ стимулирует разрыв граафова пузырька и овуляцию (Hurk Van Den R., 1994).
ЛГ является основным регулятором синтеза стероидов в яичниках. Рецеп-торы к ЛГ локализуются на лютеальных клетках, и влияние ЛГ опосредуется через стимуляцию аденилатциклазы и внутриклеточного повышения уровня цАМФ, который непосредственно или через посредников (протеинкиназа и др.) активирует ферменты, участвующие в биосинтезе прогестерона. Под влиянием ЛГ в яичниках увеличивается количество холестерина, необходи-мого для синтеза гормонов. Одновременно повышается активность фермен-тов семейства цитохромов Р450, отщепляющего боковую цепь в молекуле холестерина. При более длительном влиянии ЛГ стимулирует экспрессию и синтез других ферментов (ЗВ-гидроксистероидная дегидрогеназа, 17а-гидроксилаза), участвующих в синтезе прогестерона и других стероидов. Таким образом, в желтом теле под влиянием ЛГ усиливаются процессы стеро-идогенеза на участке конверсии холестерина в прегнанолон (Yen S., 1999).
Регуляция секреции гонадотропинов обеспечивается за счёт "короткой" и "ультракороткой" цепей обратной связи. Так, повышение уровня ЛГ и ФСГ приводит к торможению их синтеза и высвобождения, а повышенная концен-трация гонадолиберина в гипоталамусе угнетает его синтез и высвобождение в портальную систему гипофиза (Sopelak V.M., 1997).
На высвобождение гонадолиберина оказывают влияние также катехолами-ны: дофамин, адреналин и норадреналин. Адреналин и норадреналин стиму-лируют высвобождение гонадолиберина, тогда как дофамин оказывает такое же действие только у животных, которым предварительно вводились стероид-ные гормоны. Холецистокинин, гастрин, нейротензин, опиоиды и соматоста-тин угнетают высвобождение гонадолиберина (Yen S., 1999).
Адренокортикотропный гормон оказывает стимулирующее действие на кору надпочечников. За счет повышения синтеза белка (цАМФ-зависимая ак-тивация) происходит гиперплазия коркового вещества надпочечников. АКТГ усиливает синтез холестерина и скорость образования прегнанолона из холе-стерина. В большей степени его влияние выражено на пучковую зону, что приводит к увеличению образования глюкокортикоидов, в меньшей — на клубочковую и сетчатую зоны, поэтому он не оказывает значительного воз-действия на продукцию минералокортикоидов и половых гормонов.
Вненадпочечниковые эффекты АКТГ заключаются в стимуляции липолиза (мобилизует жиры из жировых депо и способствует окислению жиров), увели-чении секреции инсулина и соматотропина, накоплении гликогена в клетках мышечной ткани, гипогликемии, что связано с повышенной секрецией инсули-на, усилении пигментации за счет действия на пигментные клетки меланофоры.
Соматотропный гормон принимает участие в регуляции процессов роста и физического развития, оказывая стимулирующее действие на образование бел-ков в организме, синтеза РНК и транспорта аминокислот из крови в клетки.
Основная биологическая роль пролактина — рост молочных желез и регу-ляция лактации. Это осуществляется путём стимуляции синтеза белка — лактальбумина, жиров и углеводов молока. Пролактин регулирует также обра-зование желтого тела и выработку им прогестерона, влияет на водно-солевой обмен организма, задерживая воду и натрий в организме, усиливает эффекты альдостерона и вазопрессина, повышает образование жира из углеводов.
Гормоны задней доли гипофиза образуются в гипоталамусе. В нейрогипо-физе происходит их накопление. В клетках супраоптического и паравентри-кулярного ядер гипоталамуса осуществляется синтез окситоцина и антидиу-ретического гормона. Синтезированные гормоны путем аксонального транс-порта с помощью белка-переносчика нейрофизина по гипоталамо-гипофизар-ному тракту транспортируются в заднюю долю гипофиза. Здесь происходит депонирование гормонов и в дальнейшем их выделение в кровь.
Антидиуретический гормон (АДГ), или вазопрессин, осуществляет в орга-низме две основные функции. Антидиуретическое его действие заключается в стимуляции реабсорбции воды в дистальном отделе нефрона. Это действие осуществляется благодаря взаимодействию гормона со специфическими ре-цепторами, что приводит к повышению проницаемости стенки канальцев, ее реабсорбции и концентрированию мочи. Повышение реабсорбции воды при этом происходит также за счёт активация гиалуронидазы в клетках канальцев, что приводит к усилению деполимеризации гиалуроновой кислоты, в резуль-тате чего и увеличивается объем циркулирующей жидкости.
В больших дозах (фармакологических) АДГ суживает артериолы, в резуль-тате чего повышается артериальное давление. Поэтому его также называют вазопрессином. При его физиологических концентрациях в крови это дейст-вие не имеет существенного значения. Увеличение выброса АДГ, которое происходит при кровопотере, болевом шоке, осуществляет сужение сосудов, имеющее в этих случаях адаптивное значение.
Повышение выработки АДГ происходит при уменьшении объема внекле-точной и внутриклеточной жидкости, снижении артериального давления, уве-личении осмотического давления крови, при активации ренин-ангиотензино-вой и симпатической нервной системы.
Окситоцин избирательно действует на гладкую мускулатуру матки, вы-зывая ее сокращения при родах. Этот процесс осуществляется за счёт связы-вания со специальными окситоциновыми рецепторами, расположенными на поверхностной мембране клеток. Под влиянием высоких концентраций эст-рогенов резко возрастает чувствительность рецепторов к окситоцину, чем и объясняется повышение сократительной активности матки перед родами.
Участие окситоцина в процессе лактации заключается в усилении сокра-щения миоэпителиальных клеток молочных желез, за счёт чего увеличивает-ся выделение молока. Увеличение секреции окситоцина, в свою очередь, про-исходит под влиянием импульсов от рецепторов шейки матки, а также меха-норецепторов сосков грудной железы при кормлении грудью.
Следующий уровень репродуктивной системы — яичники, в которых происходит стероидо- и фолликулогенез в ответ на циклическую секрецию гонадотропинов и под влиянием факторов роста (ФР).
Яичник — парный орган женской репродуктивной системы и одновременно железа внутренней секреции. Яичник состоит из двух слоев: коркового вещества, покрытого белочной оболочкой, и мозгового вещества. Отдельно рассматривает-ся участок ворот яичника, лишенный тека-лютеиновых клеток стромы, содержа-щий зернистые клетки, которые отвечают за выработку яичниковых андрогенов.
Корковое вещество образовано фолликулами различной степени зрелости (от примордиальных до атрезирующихся), расположенными в соединитель-нотканной строме.
Процесс фолликулогенеза происходит в яичнике непрерывно и регулиру-ется гонадотропинами путём взаимодействия с яичниковыми рецепторами (Sopelak V.M., 1997).
Одновременно в каждом яичнике выявляется несколько десятков фоллику-лов, находящихся в различных стадиях роста и созревания. Общее число фол-ликулов к рождению составляет около 2 млн. Их число сокращается в 8-10 раз к моменту установления менструального цикла, не превышая 30-40 тыс. Толь-ко около 10% фолликулов проходит полный цикл развития от премордиаль-ного до овуляторного и превращается в желтое тело. Остальные подвергают-ся атрезии и обратному развитию (Hurk Van Den R. et al., 1994).
В ходе превращения первичного фолликула в зрелый завершается первое деление мейоза, в результате чего высвобождается однонаправительное (по-лярное) тельце и образуется овоцит. Прозрачная оболочка достигает макси-мального развития, превращаясь в лучистый венец, покрытый 1-2 слоями ха-отично лежащих фолликулярных клеток. В фолликуле образуется полость, которая достигает максимального размера перед овуляцией. Слой фоллику-лярных клеток под действием факторов роста кровеносных сосудов стромы превращается в два слоя: внутреннюю и наружную теку фолликула. Дальней-шее увеличение количества фолликулярной жидкости ведет к переполнению полости фолликула и его разрыву — овуляции. После овуляции овоцит, окру-женный лучистым венцом, попадает из брюшной полости в воронку маточной трубы и далее — в ее просвет. Здесь завершается второе деление мейоза и об-разуется зрелая яйцеклетка, готовая к оплодотворению (Yen S., 1999).
Овариальный цикл состоит из двух фаз — фолликулярной и лютеино- вой, которые разделены овуляцией и менструацией.
В фолликулярной фазе под влиянием ФСГ, секретируемым гипофизом, совместно с различными факторами роста происходит стимуляция роста и развития одного или нескольких примордиальных фолликулов, а также диф-ференцировка и пролиферация клеток гранулезы. ФСГ также потенцирует ак-тивность 17-(3-гидроксистероидной дегидрогеназы и ароматазы, которые не-обходимы для образования эстрадиола в клетках гранулезы через активацию цАМФ, и таким образом стимулирует процессы роста и развития первичных фолликулов, выработку эстрогенов клетками фолликулярного эпителия. Эст-радиол же, в свою очередь, повышает чувствительность клеток гранулезы к действию ФСГ. Рецепторы к ФСГ относятся к группе мембранных рецепто-ров, имеющих 7 трансмембранных фрагментов. Наряду с эстрогенами секре-тируются небольшие количества прогестерона. Из множества начинающих рост фолликулов окончательной зрелости достигнет только 1, реже — 2-3.
Предовуляторный выброс гонадотропинов определяет сам процесс овуляции. Объем фолликула быстро увеличивается параллельно с истончением стенки фолликула, связанным с повышенной активностью протеолитических фермен-тов и гиалуронидазы, выделяемых полиморфно-ядерными лейкоцитами.
Наблюдающееся в течение 2-3 дней, предшествующих овуляции, значи-тельное увеличение уровня эстрогенов обусловлено гибелью большого числа зрелых фолликулов с высвобождением фолликулярной жидкости. Высокие концентрации эстрогенов по механизму отрицательной обратной связи инги-бируют секрецию ФСГ гипофизом. Овуляторный выброс ЛГ и в меньшей степени ФСГ связан с существованием механизма положительной обратной связи сверхвысоких концентраций эстрогенов и уровня ЛГ, а также с резким падением уровня эстрадиола в течение 24 ч, предшествующих овуляции.
Нейрогормональная регуляция менструального цикла схематически пред-ставлена на рисунке 6.
Овуляция
Рисунок 6. Нейрогормональная регуляция менструального цикла
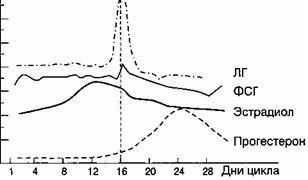
Овуляция яйцеклетки происходит лишь в присутствии ЛГ или хориони-ческого гонадотропина. Более того, ФСГ и ЛГ выступают как синергисты в период развития фолликула, и в это время тека-клетки активно секретируют эстрогены.
Механизм разрушения коллагенового слоя стенки фолликула — гормо-нально-зависимый процесс, в основе которого лежит адекватность фоллику-лярной фазы. Предовуляторный выброс ЛГ стимулирует повышение концен-трации прогестерона к моменту овуляции. Благодаря первому пику прогестерона повышается эластичность фолликулярной стенки, таким обра-зом ФСГ, ЛГ и прогестерон совместно стимулируют активность протеолити-ческих ферментов: активаторы плазминогена, секретируемые клетками гра-нулезы, способствуют образованию плазмина, плазмин вырабатывает различные коллагеназы, простагландины Е и F 2 ot способствуют вытеснению скопления клеточной массы овоцита. Для того, чтобы не произошла прежде-временная лютеинизация неовулирующего фолликула, в яичнике должно вы-рабатываться определенное количество активина (Speroff L. et al., 1994).
После овуляции отмечается резкое снижение уровня ЛГ и ФСГ в сыворотке крови. С 12-го дня второй фазы цикла отмечается 2-3-дневное повышение уровня ФСГ в крови, которое инициирует созревание нового фолликула, тогда как кон-центрация ЛГ в течение всей второй фазы цикла имеет тенденцию к снижению.
Полость совулировавшего фолликула спадается, а стенки его собираются в складки. Вследствие разрыва сосудов в момент овуляции в полость постовуля-торного фолликула происходит кровоизлияние. В центре будущего желтого те-ла появляется соединительнотканный рубец — стигма (Speroff L. et al., 1994).
Овуляторный выброс ЛГ и последующее за ним поддержание высокого уровня гормона в течение 5-7 дней активирует процесс пролиферации и желе-зистого метаморфоза клеток зернистой зоны (гранулезы) с образованием лю-теиновых клеток, т.е. наступает лютеиновая фаза (фаза желтого тела) овари-ального цикла (Erickson G.F., 2000).
Эпителиальные клетки зернистого слоя фолликула интенсивно размножа-ются и, накапливая липохромы, превращаются в лютеиновые клетки; сама оболочка обильно васкуляризуется. Стадия васкуляризации характеризуется быстрым размножением эпителиальных клеток гранулезы и интенсивным врастанием между ними капилляров. Сосуды проникают в полость постову-ляторного фолликула со стороны thecae internae в лютеиновую ткань в ради-альном направлении. Каждая клетка желтого тела богато снабжена капилля-рами. Соединительная ткань и кровеносные сосуды, достигая центральной полости, заполняют ее кровью, окутывают последнюю, ограничивая от слоя лютеиновых клеток. В желтом теле — один из самых высоких уровней крово-тока в организме человека. Формирование этой уникальной сети кровеносных сосудов заканчивается в течение 3-4 дней после овуляции и совпадает с пери-одом расцвета функции желтого тела (Bagavandoss P., 1991).
Ангиогенез состоит из трех фаз: фрагментации существующей базальной мембраны, миграции эндотелиальных клеток и их пролиферации в ответ на митогенный стимул. Ангиогенная активность находится под контролем основ-ных ростовых факторов: фактора роста фибробластов (ФРФ), эпидермального фактора роста (ЭФР), фактора роста тромбоцитов (ФРТ), инсулиноподобного фактора роста-1 (ИФР-1), а также цитокинов, таких как тумор некротического фактора (ТНФ) и интерлейкинов (ИЛ-1; ИЛ-6) (Bagavandoss P., 1991).
С этого момента желтое тело начинает продуцировать значительные количе-ства прогестерона. Прогестерон временно инактивирует положительный меха-низм обратной связи, и секреция гонадотропинов контролируется только нега-тивным влиянием зстрадиола. Это приводит к снижению уровня гонадотропинов в середине фазы желтого тела до минимальных значений (Erickson G.F., 2000).
Прогестерон, синтезируемый клетками желтого тела, ингибирует рост и развитие новых фолликулов, а также участвует в подготовке эндометрия к внедрению оплодотворенной яйцеклетки, снижает возбудимость миометрия, подавляет действие эстрогенов на эндометрий в секреторной фазе цикла, сти-мулирует развитие децидуальной ткани и рост альвеол в молочные железах. Плато сывороточной концентрации прогестерона соответствует плато рек-тальной (базальной) температуры (37,2—37,5°С), что лежит в основе одного из методов диагностики произошедшей овуляции и является критерием оцен-ки полноценности лютеиновой фазы. В основе повышения базальной темпе-ратуры лежит снижение под воздействием прогестерона периферического кровотока, что уменьшает теплопотерю. Увеличение его содержания в крови совпадает с повышением базальной температуры тела, которая является инди-катором овуляции (McDonnel D.P., 2000).
Прогестерон, являясь антагонистом эстрогенов, ограничивает их пролифера-тивный эффект в эндометрии, миометрии и эпителии влагалища, вызывая стиму-ляцию секреции железами эндометрия секрета, содержащего гликоген, уменьшая строму подслизистого слоя, т.е. вызывает характерные изменения эндометрия, необходимые для имплантации оплодотворенной яйцеклетки. Прогестерон сни-жает тонус мышц матки, вызывает их расслабление. Кроме того, прогестерон вы-зывает пролиферацию и развитие молочных желез и в период беременности спо-собствует угнетению процесса овуляции (O"Malleu B.W., Strott G.A., 1999).
Продолжительность этой фазы развития фолликула различна: если опло-дотворения не произошло, то через 10-12 дней наступает регресс менструаль-ного желтого тела, если же оплодотворенная яйцеклетка внедрилась в эндо-метрий и образующаяся бластула стала синтезировать хорионический гонадотропин (ХГ), то желтое тело становится желтым телом беременности.
Клетками гранулезы желтого тела секретируется полипептидный гормон релаксин, который принимает важное участие в период родов, вызывая релак-сацию связок таза и расслабление шейки матки, а также увеличивает синтез гликогена и задержку воды в миометрии, уменьшая при этом его сократитель-ную способность. В период обычного менструального цикла секреция его повышается сразу после пика высвобождения ЛГ и остается определяемым в период менструации. В течение беременности циркулирующий уровень ре-лаксина выше в конце I триместра по сравнению со II и III триместрами.
Если не произошло оплодотворение яйцеклетки, желтое тело переходит в ста-дию обратного развития, что сопровождается менструацией. Лютеиновые клетки подвергаются дистрофическим изменениям, уменьшаются в размерах, при этом наблюдается пикноз ядер. Соединительная ткань, врастая между распадающими-ся лютеиновыми клетками, замещает их, и желтое тело постепенно превращает-ся в гиалиновое образование — белое тело {corpus albicans) (Sopelak V.M., 1997).
С точки зрения гормональной регуляции, период регресса желтого тела харак-теризуется выраженным снижением уровней прогестерона, эстрадиола и ингиби-на А. Падение уровня ингибина А устраняет его блокирующий эффект на гипо-физ и секрецию ФСГ. В то же время прогрессирующее снижение концентрации эстрадиола и прогестерона способствует быстрому повышению частоты секре-ции Гн-РГ, и гипофиз освобождается от торможения отрицательной обратной связи. Снижение уровней ингибина А и эстрадиола, а также возрастание частоты импульсов секреции Гн-РГ обеспечивают преобладание секреции ФСГ над ЛГ. В ответ на повышение уровня ФСГ окончательно формируется пул антральных фолликулов, из которых в дальнейшем будет выбран доминантный фолликул. Простагландин F 2 a, окситоцин, цитокины, пролактин и радикалы 0 2 обладают лютеолитическим эффектом, что может быть основанием для развития недоста-точности желтого тела при наличии воспалительного процесса в придатках.
Длительность овариального (менструального) цикла в норме варьирует от 21 до 35 дней.
Менструация происходит на фоне регресса желтого тела. К ее окончанию уровни эстрогенов и прогестерона достигают своего минимума. На этом фоне происходят активация тонического центра гипоталамуса и гипофиза и повышение секреции преимущественно ФСГ, активирующего рост фолликулов. Повышение уровня эс-традиола ведет к стимуляции пролиферативных процессов в базальном слое эндо-метрия, что обеспечивает адекватную регенерацию эндометрия (рис. 7).
Рисунок 7. Звенья регуляции нормального менструального цикла (Sopelak V., 1997)
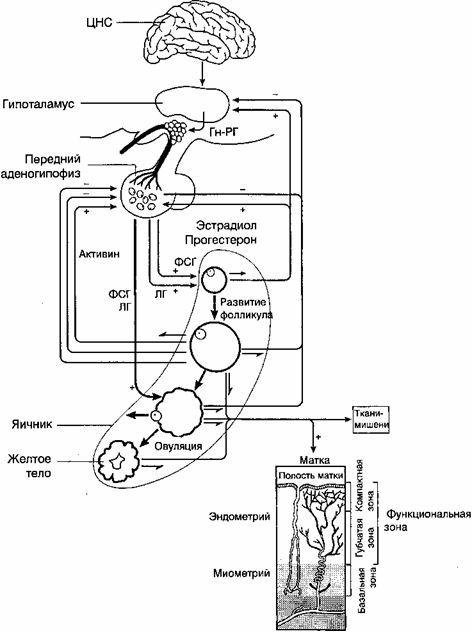
Яичниковый стероидогенез проходит в клетках эпителия, выстилающих по-лость фолликула, в клетках внутренней теки и значительно меньше в строме. Фолликулярные эпителиоциты, стромальная и тека-ткань синтезируют прогесте-рон, тестостерон, дегидротестостерон, эстрон и эстрадиол (Erickson G.F., 2000).
Эстрогены представлены эстрадиолом, эстроном и эстриолом. Биологичес-ки наиболее активен эстрадиол, 95% которого образуется в фолликуле, и уро-вень его в крови является показателем созревания фолликула. Эстрадиол (Е 2) секретируется преимущественно клетками гранулезы, а также, в меньших ко-личествах, желтым телом. Эстрон (Е,) образуется путем периферической аро-матизации эстрадиола. Основным источником эстриола (Е 3) является гидро-ксилирование эстрадиола и эстрона в печени (O"Malleu B.W., Strott G.A., 1999).
Секретируемые в кровь эстрогены конъюгируются сексстероидсвязываю-щим глобулином (СССГ) и в меньшей степени альбуминами крови. СССГ иначе называется эстрадиол-тестостерон-связывающим глобулином. Уже са-мо название указывает на повышенное сродство этого белка к андрогенам. Уровень глобулина, связывающего половые гормоны, в сыворотке крови жен-щин почти в 2 раза выше по сравнению с его концентрацией в крови мужчин. Эстрогены и их метаболиты конъюгируются в печени с глюкуроновой и сер-ной кислотами и экскретируются с желчью и мочой (McDonnel D.P., 2000).
Кроме уже упомянутого влияния на половые органы, гипофиз и гипотала-мус, эстрогены обладают анаболическим свойством, усиливают обмен костной ткани и ускоряют созревание костей скелета, с чем связано прекращение рос-та при наступлении полового созревания, с одной стороны, и развитие юве-нильного остеопороза у девочек при задержке полового развития — с другой.
В больших дозах эстрогены способствуют задержке натрия и воды в орга-низме вплоть до развития отеков. Влияют также на обмен липидов, снижая уровень холестерина в крови.
Прогестерон секретируется желтым телом, а также корой надпочечников и яичками, где используется как предшественник для биосинтеза кортикостерои-дов и андрогенов. Прогестагены и глюкокортикоиды имеют сходную химичес-кую структуру, поэтому прогестероновые и глюкокортикоидные рецепторы имеют перекрёстно-связывающие свойства. В сыворотке крови прогестерон свя-зывается транскортином, который, как известно, связывает и глюкокортикоиды. По данным некоторых исследований, способность прогестерона связываться транскортином даже превышает соответствующую у кортикостероидов. В пече-ни прогестерон связывается глюкуроновой кислотой и в конъюгированном состоянии экскретируются с мочой (McDonnel D.P., 2000). Однако более подробно влияние эстрогенов и прогестерона на органы-мишени изложено в разделе "Принципы применения половых стероидных гормонов в клинической практике и их системные эффекты".
Андрогены у женщин секретируются клетками стромы яичников, главным образом в виде андростендиона, причем в надпочечниках его образуется в 3 раза больше, чем в яичниках. Андростендион в периферических тканях конвертируется в тестостерон. В яичниках образуется в незначительных количествах также тестостерон, дигидротестостерон, дегидроэпиандросте-рон. Приблизительно около 1/4 тестостерона, который секретируется в орга-низме женщины, образуется в яичниках. Остальное его количество секретиру-ется надпочечниками или образуется в тканях на периферии путем конверсии из андростендиона (McDonnel D.P., 2000).
Биологическое действие стероидов в тканях-мишенях связано с наличием в них специфических рецепторов (рис. 8). Стероиды путем диффузии прохо-дят мембрану клетки и в цитоплазме связываются со специфическими рецеп-торами. Стероидные рецепторы представляют собой относительно большие белки с высокой связывающей способностью к определённым гормонам. Од-нако возможно связывание этих рецепторов с другими стероидами данной группы (например, с синтетическими агонистами и антагонистами). Цито-плазматические рецепторы присутствуют не во всех, а только в клетках тка-ней, чувствительных к данному виду гормона. Рецепторно-стероидный ком-плекс, образование которого зависит от нескольких факторов, включая температуру, перемещается в ядро, где на хроматине имеются специальные участки, связывающие эти комплексы. Комплекс рецептор-стероид становит-ся активированным, после чего возможно его связывание с акцепторным ядерным белком, расположенным на ДНК. Последнее взаимодействие ведет к синтезу большого количества специфических РНК и соответствующих бел-ков, росту и развитию соответствущих органов (молочные железы, матка и др.) и тканей (O"Malleu B.W., Strott G.A., 1999).
|
|
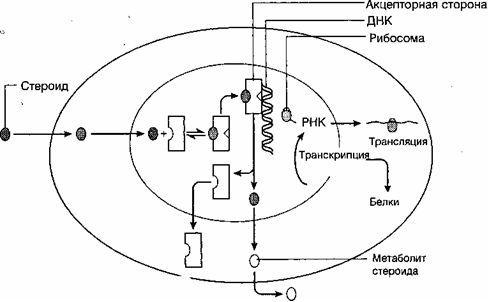
Количество молекул рецепторов для различных стероидных гормонов ко-леблется от 5000 до 20 000 на клетку. Рецепторы к эстрогенам связывают многие естественные и синтетические эстрогенные стероиды с одинаковой аф-финностью. Считается, что рецепторы к эстрогенам и прогестерону представ-ляют собой две субъединицы, каждая из которых связывает молекулу гормо-на, о чём более подробно изложено в разделе клинической практики.
Каждая из субъединиц а и Р взаимодействует с хроматином и обеспечива-ет дальнейшую активацию специфических генов и РНК-полимераз. Биологическое действие гормона связано не только с количественными ко-лебаниями его в сыворотке крови, но и с состоянием рецепторного звена, при-чем количество рецепторов подвержено значительным колебаниям.
|
Рисунок 8. Механизм действия стероидных гормонов на ткани-мишени (Cowan B.D., 1997) |
Экспери-ментальные исследования показали, что у новорожденных крыс ткани-мишени содержат незначительное количество рецепторов к эстроге-нам. На 10-й день жизни количество рецепторов возрастает, и после этого срока введение экзогенных эстрогенов вызывает их увеличение. Эстрогены стимулируют образование рецепторов не только к эстрогенам, но и к прогес-терону. Количество рецепторов не только зависит от уровня циркулирующе-го в крови гормона, но и находится под генетическим контролем. Так, полное отсутствие рецепторов к андрогенам наблюдается при синдроме тестикуляр-ной феминизации (McDonnel D.P., 1999).
|
|
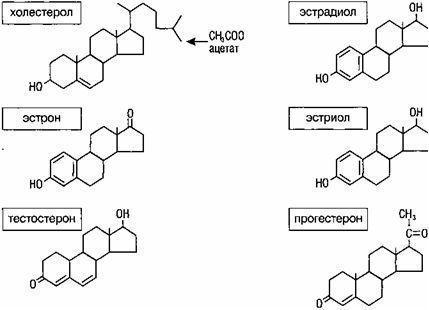
Анализ химической структуры основных половых стероидных гормонов показывает, что все они являются производными прогестерона, причем эст-рогены между собой отличаются лишь количеством имеющихся в их структу-ре гидроксирадикалов (рис. 9).
Субстанцией для всех стероидных гормонов является холестерин липопро-теидов низкой плотности (ЛПНП). В стероидогенезе участвуют гонадотропины (ФСГ и ЛГ), а также ферментные системы (ароматазы). Вначале образуется прегнанолон в результате отщепления боковой цепи холестерина. В дальней-шем возможно два пути метаболических превращений прегнанолона, заканчи-вающихся образованием тестостерона, получивших по положению двойной не-насыщенной связи в образующихся соединениях названия м - и л5 -путей метаболизма.
Преимущественное образование половых стероидов происходит по Л5 -пути.
|
Рисунок 9. Химическая структура стероидных гормонов (Sopelak V., 1997) |
В его ходе образуются последовательно 17а-гидроксипрегнанолон, дегидроэпиандростерон (ДГЭА), андростендион.
По Л4 -пути образуются прогестерон, 17а-гидроксипрогестерон, андростендион. Замыкает оба пути А4,5 -изомераза. Далее происходит ароматизация тестостерона или андростендиона с образованием, соответственно, эстрадиола или эстрона (рис. 10).
Примечание: ГСД — Зр-гидроксистероиддегидрогеназа, ДОК — дезоксикортикостерон
Рисунок 10. Биосинтез стероидов (Cowan B.D., 1997)
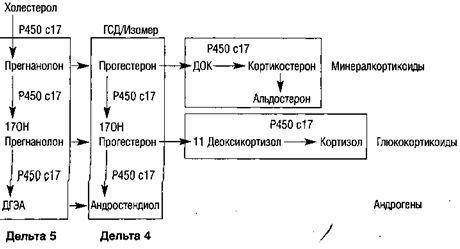
Большинство стероидогенных ферментов, превращающих холестерол в предшественники и в биологически активные стероиды, входят в группу ци-тохромов Р450. Цитохром Р450 — это генерический термин для многих окис-лительных ферментов (Bryan D., 1997). Существует около 200 типов цитохро-мов, из них в процесс стероидогенеза вовлечены пять (табл. 2).
Ферменты Р450, участвующие в процессе стероидогенеза
|
Р 450 - новое название |
Ароматаза |
Таким образом, репродуктивная система — это единая целостная система, все звенья которой взаимосвязаны как механизмом прямой, так и обратной связи.
Медицинский центр "Семья" в Ростове-на-Дону – это многопрофильная клиника для взрослых и детей